 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Zootecnia Tropical, Vol. 23, No. 2, 2005, pp. 121-139 Identificación botánica y producción de frutos en un bosque deciduo del asentamiento Las Peñitas, al sur del estado Aragua, Venezuela Botanical identification and fruit production in a deciduous forest at Las Peñitas settlement, south of Aragua state, Venezuela José Valero1, Miguel Benezra1*, Selina Camacaro1, Luís Chong2 y Orlando Guenni2 1Instituto
de Producción Animal, Facultad de Agronomía, Universidad
Central de Venezuela. Maracay, estado Aragua. Venezuela. Code Number: zt05010 RESUMEN Se colectaron muestras vegetales con el objeto de identificar botánicamente las especies leñosas presentes en un bosque deciduo, así como su frecuencia, densidad y el grado de diversidad dentro del bosque. El bosque en estudio se dividió en cuatro áreas diferentes: bosque no intervenido, bosque no intervenido recientemente, bosque parcialmente intervenido recientemente y bosque totalmente intervenido recientemente y en cada una se establecieron 3 transectas de 100 m x 5 m. La frecuencia y densidad se determinaron por el número de individuos de cada especie presentes en cada transecta y el grado de diversidad se estimó mediante los índices de Simpson y de Shannon. Se encontraron 32 especies botánicas diferentes, predominando la familia Leguminoseae con un 37% de participación y otras 14 familias que representaron el otro 63%. Se evidenciaron diferencias en la cantidad y la frecuencia de las especies entre cada una de las áreas fisonómicas, así como diferencias entre especies en la misma área. El área con mayor densidad de especies fue el bosque no intervenido (2.780 plantas/ha). El grado de diversidad evaluado por los índices de Simpson y de Shannon resultó mayor (P<0,05) para el bosque no intervenido (15,1 y 4,01, respectivamente). Así, aunque todas estas comunidades tienen potencial para la alimentación animal, el bosque no intervenido parece tener el mayor potencial para la alimentación de rumiantes, por presentar mejores indicadores botánicos. Palabras clave: bosque deciduo, producción de frutos, caracterización botánica, frecuencia, diversidad. SUMMARY Herbarium specimen were collected in order to identify the woody species present in a deciduous forest, as well as their frequency, density and the degree of diversity of the forest. The studied forest was divided in four different areas: no intervened forest, forest no recently intervened, forest partially intervened recently, and forest completely intervened recently, and in each area 3 experimental plots of 100 m x 5 m were set along topography gradients. The frequency and density were determined by the number of individuals of each species in the plot, and the degree of diversity was assumed by the Simpson and Shannon’s indexes. There were 32 different botanical species, predominating the Leguminoseae family with 37% and 14 other families that represented 63%. Differences were found in the quantity and frequency of the species among the physiognomic areas, as well as differences among species in the same area. The area with higher species density was the no intervened forest (2780 plants/ha). The degree of diversity evaluated by the Simpson and Shannon´s indexes was higher (P<0.05) for the not intervened forest (15.1 and 4.01, respectively). Although all these communities have the potential for animal feeding, the no intervened forest seems to have the best potential for this purpose because of better botanical indicators. Key words: deciduous forest, fruit production, botanical characterization, frequency, diversity. INTRODUCCIÓN La mayor limitante de los sistemas ganaderos en Venezuela es la baja disponibilidad de forrajes en la época seca, debido a que estos disminuyen dramáticamente su crecimiento. El uso racional del bosque deciduo puede ser una alternativa para la alimentación de rumiantes durante este periodo, por su considerable producción de hojarasca y frutos (Baldizán y Chacón, 2001). El estudio realizado por Molina (1998) caracterizó la zona de estudio desde el punto de vista fisonómico, encontrando que predominan los bosques bajos, los cuales representan el bosque maduro para este tipo de condiciones ecológicas, no encontrándose presencia de bosques muy altos en la zona. Entre las especies presentes en estos bosques están: Birote (Caesalpinia mollis), Acapro (Tabebuia spectabilis), Barbasco (Pscidia guaricencis), Melero (Combretum fruticosum), Roble (Platymisciun pinnatum), Guasimo (Guazuma tomentosa), Cují (Mimosa tenuiflora), Pardillo (Cordia sp.), Drago (Pterocarpus podocarpus) y Jobo (Spondias mombín). Esto evidenció la diversidad de estos bosques y su elevado potencial forrajero. En los sistemas silvopastoriles se han identificado una gran diversidad de especies arbóreas con alto potencial para la alimentación de rumiantes. En general, se acepta que el mayor potencial para la alimentación de estos animales, se encuentra en las leguminosas. Sin embargo, muchas especies distintas a las leguminosas son también potencialmente utilizable por los animales, dependiendo del genotipo y de las condiciones medioambientales y socioeconómicas locales (Giraldo, 1994). En estos ecosistemas, los animales actúan como consumidores de material vegetal, básicamente frutos, hojas y ramas tiernas, descartando el material lignificado. La biomasa es empleada para satisfacer sus necesidades de mantenimiento y producción, siendo los rumiantes las especies animales más eficientes en la conversión de esta biomasa foliar, gracias a las peculiaridades de su aparato digestivo y a la asociación simbiótica en el rumen con diversos microorganismos (Giraldo y Vélez, 1993). El valor nutricional de las especies varía entre los diferentes componentes de la biomasa arbórea. Las hojas presentan mayores concentraciones de nutrientes que las ramas y tallos; la variación también se ha relacionado con la especie, con la edad y con la relación tallo/hoja del árbol. Estos árboles forrajeros pueden tener contenidos de proteína cruda superiores al 12%. En 35 especies arbóreas evaluadas, 20 de ellas mostraron mas de 60% de digestibilidad in vitro de la materia seca (Benavides, 1993). En este estudio se evaluó la composición botánica, frecuencia, densidad de las especies vegetales y el grado de diversidad de un bosque deciduo con el fin de inferir el potencial de esta comunidad vegetal en la alimentación de rumiantes bajo sistemas silvopastoriles ubicados en la zona de colinas del sur del estado Aragua. MATERIALES Y MÉTODOS El trabajo se realizó en la parroquia Las Peñitas del municipio Urdaneta, al sur del estado Aragua, en una unidad de producción del asentamiento campesino Las Peñitas. La zona en estudio limita por el norte con la parroquia Taguay, por el sur y el este con los municipios Chaguaramas y Monagas respectivamente del estado Guárico y por el oeste con la parroquia de Barbacoas. Las coordenadas geográficas aproximadas son entre 9°20`59” y 9°21`15” N y entre 66°35`16” y 66°35`33” E (Molina, 1998). La zona presenta clima tropical seco con temperaturas máximas medias que alcanzan 33°C y temperaturas mínimas medias entre 22 y 24°C, sin registrarse variaciones significativas entre años, siendo la precipitación promedio anual de 800 a 1.000 mm (Moreno et al., 1995). La distribución de las lluvias se concentra en 5 ó 6 meses (mayo a septiembre), existiendo una alta evaporación, la cual supera los 2.400 mm, que provoca una carencia de agua el resto del año. Esta marcada estacionalidad de la precipitación conlleva a una producción agrícola estacional y una oferta forrajera intermitente En la zona se presentan una gran diversidad de suelos que se caracterizan por su distribución irregular. En la altiplanicie se observa dominancia de suelos clasificados como Vertisoles (60%), relacionados con relieves ondulados, en asociación con suelos de tipo Alfisoles (20%), ubicados en zonas fuertemente onduladas a onduladas. En pequeña proporción y en relieves ondulados y moderadamente erodados se presentan áreas con suelos tipo Ultisoles (Molina, 1998). El bosque deciduo se inspeccionó visualmente por medio de visitas de campo y se dividió en cuatro áreas diferentes tomando en cuenta sus aspectos fisonómicos y la información histórica suministrada por el propietario, relativa a la deforestación realizada y el tiempo aproximado de la misma. Estas cuatro áreas se denominaron: bosque no intervenido (BNI), el cual no ha sido deforestado en los últimos 20 años, bosque no intervenido recientemente (BNIR), cuando la ultima deforestación fue ligera, selectiva y hace mas de 10 años, bosque parcialmente intervenido recientemente (BPIR), cuando la intervención tiene menos de 10 años y en forma selectiva y bosque totalmente intervenido recientemente (BTIR), cuando la deforestación fue total y hace menos de 10 años. La superficie aproximada de cada una de las parcelas delimitadas para el estudio fue de 30.000 m2 para el BNI y 25.000 m2 para el BNIR, BPIR y BTIR, para un total de 105.000 m2. Utilizando un diseño completamente al azar, en cada una de estas parcelas se establecieron 3 transectas de 100 m de largo y 5 m de ancho (500 m2 c/u), las cuales fueron analizadas como repeticiones. Las muestras de herbario para la identificación botánica se colectaron de todas las especies leñosas presentes en el recorrido de las transectas, las cuales se prensaron y se secaron y fueron llevadas al Herbario del Instituto de Botánica de la Facultad de Agronomía de la Universidad Central de Venezuela, para su identificación. Se determinó la frecuencia de las especies leñosas (árboles, arbustos y trepadoras) de acuerdo al número de individuos de cada especie presente en el área de la transecta, expresándola en forma absoluta (número de individuos por transecta) y relativa (% de frecuencia de la especie con respecto al total de plantas en la transecta). Utilizando los valores de la frecuencia absoluta por especie en las transectas y conociendo el área de muestreo, se estimó la densidad de cada una de ellas, expresándola como plantas/ha (Smith, 1996). Se evaluó también la producción de frutos por planta en las cuatro especies que por experiencia local señalada por el propietario son mas consumidas por los rumiantes (Caesalpinia granadillo, Acacia macracanta, Caesalpinia coriaria y Senna atomaria). Para ello se colectó el total de los frutos caídos al suelo durante la época seca en 5 plantas por especie. La producción de frutos por hectárea se estimó usando los valores anteriores y la densidad de la especie en las diferentes áreas fisonómicas. El grado de diversidad de la cobertura vegetal leñosa se estimó mediante el índice de Simpson (Simpson, 1949) y la fórmula de Shannon (Shannon y Wiener, 1963). El índice de diversidad de Simpson se calculó por medio de la siguiente ecuación:
donde:
El índice de Shannon se calculó por la siguiente ecuación:
donde:
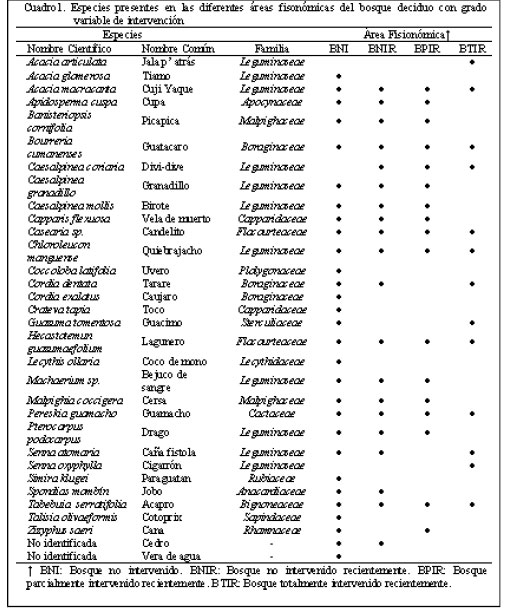
Los resultados obtenidos por los dos índices anteriores fueron analizados por el método de los cuadrados mínimos en un análisis de varianza, utilizando el Programa Statistix for Windows. Para la comparación de medias entre áreas muestreadas se utilizó la prueba de Tukey. RESULTADOS Y DISCUSIÓN Se observaron diferencias importantes en el aspecto fisonómico entre el BNI y el BTIR entre si y con las otras áreas, no resultando evidentes las diferencias entre el BNIR y el BPIR. Estas dos últimas se separaron solo en función de la reseña histórica señalada por el productor. En el BNI se observaron árboles de gran tamaño con copas densas, así como una proporción de plantas jóvenes relativamente menor, presentándose una densidad menor de la vegetación en los estratos más bajos. En el BTIR no se observaron árboles de gran tamaño, pero la cobertura de la vegetación fue mayor, así como el sotobosque más desarrollado. Los BNIR y BPIR mostraron un aspecto similar entre ellos con características intermedias a las descritas anteriormente. Se identificaron 15 familias, representando la familia Leguminoseae el 37% del total de las familias presentes; las otras 14 familias identificadas variaron entre 3 y 10%. Pezo (1994) señala que la familia Leguminoseae es importante en la alimentación animal, cuyos árboles y arbustos además tienen naturaleza multipropósito. Estas plantas por muchos años han jugado un papel preponderante en los sistemas agroforestales y silvopastoriles presentes en el trópico, incluso lejos de sus regiones de origen. Clavero (1996) indica que el uso de leguminosas arbóreas con alto potencial productivo y valor nutritivo, se presenta como una solución económicamente viable y socialmente aceptada para incrementar la productividad animal en las regiones tropicales. No obstante, Guelmes (1996) reportó que existen una gran cantidad de especies arbóreas que no son leguminosas, pero que han sido identificadas igualmente como útiles para la alimentación de rumiantes y de monogástricos. Por otro lado, este bosque se podría caracterizar como una comunidad vegetal con diversidad botánica y multiestratificada, lo cual se podría relacionar con mejores cualidades productivas y por lo tanto, con mayores posibilidades para el desarrollo de una variedad de sistemas de producción animal. Igualmente, la estratificación de los recursos disponibles pudiera propiciar una menor incidencia de la alteración del bosque como consecuencia de la deforestación para establecer nuevas comunidades vegetales (Baldizán y Chacón, 2001). Esto coincide con la opinión de Plaza y Sepúlveda (1996), quienes señalaron que los sistemas productivos deben estar diseñados sobre la base de la conservación de los recursos naturales y productividades sustentables. Se observaron diferencias visuales importantes en el aspecto fisonómico entre el BNI y el BTIR y con las otras áreas, no resultando evidentes las diferencias entre el BNIR y el BPIR, las cuales sólo fue posible separar en función de la reseña histórica señalada por el productor. En el BNI se observaron árboles de gran tamaño con copas densas, así como una proporción de plantas jóvenes relativamente menor, presentándose una densidad menor de la vegetación en los estratos más bajos. En el BTIR no se observaron árboles de gran tamaño, la densidad de la vegetación fue mayor, así como un sotobosque más desarrollado. Los BNIR y BPIR mostraron un aspecto similar entre ellos con características intermedias a las descritas anteriormente. En el Cuadro 1 se presentan las 32 especies botánicas identificadas y las familias a las cuales pertenecen, en las cuatro áreas bajo estudio. Un total de 7 especies botánicas estuvieron presentes en todas las áreas evaluadas, persistiendo por lo tanto a la intervención del bosque. Especies como Hecatostemun guazumaefolium, Pereskia guamacho y Tabebuia serratifolia, entre otras, parecieran tener una mayor resistencia natural, al tolerar mejor los cambios producidos en el ecosistema como consecuencia de la deforestación y la presencia de los animales. Así mismo, 11 especies se encontraron en 3 de las 4 áreas fisonómicas. De este total, 8 especies fueron encontradas en el BNI, BNIR y BPIR. Las otras 3 especies fueron encontradas también en el BNI y en el BNIR, pero a diferencia del grupo anterior fueron encontradas igualmente en el BTIR, con lo cual se puede inferir que dichas especies también son relativamente persistentes a un nivel alto de intervención. Se encontraron 4 especies presentes solo en 2 de las áreas estudiadas, distribuyéndose de forma irregular. Las especies Spondias mombin y “cedro” (no identificada), se encontraron en el BNI y el BNIR, mientras que Guazuma tomentosa se encontró solo en el BNI y el BTIR. La especie Zizyphus saeri fue encontrada solo en el BNI y el BPIR.
La causa de esta aparente variabilidad en la ubicación de estas especies no está determinada, pero es probable que estas encuentren condiciones favorables en algunas de las áreas para su desarrollo y no en otras. El último grupo, representado por aquellas especies presentes en una sola de las áreas, se encuentra conformado por 10 especies. De ellas, 8 (Cuadro 1) se encontraron presentes solamente en el BNI lo que pareciera indicar que dichas especies no persisten en la medida que el bosque primario es intervenido. Esto se debe, posiblemente, a una menor capacidad de competir en comunidades en sucesión, lo cual implicaría una perdida neta de biodiversidad, de frutos y de recursos forrajeros que pudieran ser utilizados en la alimentación de los rebaños bovinos, en aquellos bosques que han sido alterados por el hombre. Las otras 2 especies (Acacia articulata y Senna oxyphylla), se encontraron sólo en el BTIR, lo que permite inferir que su aparición es producto de la intervención y que la deforestación de los bosques contribuiría a su mayor crecimiento poblacional. En el Cuadro 2 se observa la frecuencia relativa y absoluta de las 29 especies encontradas en el BNI, evidenciándose resultados variables entre ellas. Las especies Hecatostemun guazuamefolium, Casearia sp., Guazuma tomentosa y Simira kluguei presentaron valores de frecuencia relativa de 11, 9, 12 y 13%, respectivamente. Veintiún (21) especies presentaron valores entre 1 y 7% y las especies Pterocarpus podocarpus, Crateva tapia, Spondias mombin y Banisteriopsis cornifolia mostraron una frecuencia muy baja, con valores menores al 1%. Algunas de estas especies fueron reportadas como plantas consumidas por los ovinos por Baldizán y Chacòn (2001), quienes indicaron que las plantas más consumidas por los caprinos en el bosque seco caducifolio fueron las hojas del árbol Bourreria cumanensis y del arbusto Senna oxyphyla. Los frutos secos predilectos, según los mismos autores, fueron de las especies Caesalpinea coriaria y Caesalpinea granadillo, mientras que los frutos pulposos predilectos fueron los de Spondias mombin y Copernicia tectorum. Todas estas especies a excepción de la palma llanera se encontraron en el BNI y en otras áreas fisonómicas del bosque deciduo en estudio. Para el bosque no intervenido recientemente (BNIR), los valores de frecuencia de las 20 especies encontradas se presentan en el Cuadro 3. Se observó una menor cantidad de especies y una mayor concentración de la frecuencia relativa en un menor número de ellas. Las especies Apidosperma cuspa y Casearia sp., representaron alrededor del 60% del total de las plantas presentes en este sector del bosque deciduo. El otro 40% recogió a las 18 especies restantes. Es importante resaltar que en la disminución del número de especies presentes se podrían conjugar una serie de factores, que probablemente imposibiliten la sobrevivencia de algunas de ellas, lo cual traería como consecuencia su desaparición. Dichos factores pudieran estar relacionados de nuevo con la intervención y deforestación. Baldizán y Chacón (2001) señalan que el factor antrópico, especialmente el ejercido en las actividades ganaderas como el desmonte, quema y sobrepastoreo, ha influido negativamente en el ecosistema, provocando la destrucción de la vegetación clímax que anteriormente estaba dominada por selvas o bosques deciduos más o menos cerrados o densos.
En el Cuadro 4 se presentan los valores obtenidos de frecuencia relativa y absoluta en el bosque parcialmente intervenido recientemente (BPIR). Se observó que la intervención del bosque, producto de la deforestación, disminuyó aún más la cantidad de especies botánicas presentes en el área. En este caso, sólo se observaron 17 especies y la distribución se concentró mayormente en Apidosperma cuspa (28,7%) y Tabebuia serratifolia (15,5%); las otras 15 especies representaron 55% de la frecuencia relativa, resaltando entre ellas 4 especies con valores cercanos al 10% (Caesalpinea granadillo, Chloroleucon manguense, Pereskia guamacho y Casearia sp.). Veillon (1995), en estudios realizados en un bosque deciduo, encontró que las especies con mayor proporción de número de árboles son Cereus spp. y Pereskia spp. (Cactaceae) con 8% y el Jobo (Spondias mombin) con 3%.
Al comparar las distintas áreas fisonómicas (BNI, BNIR, BPIR, BTIR) del bosque deciduo en estudio, se observó que existen diferencias importantes (P<0,05) en la cantidad de especies leñosas presentes. En el BNI se observó el mayor número de especies (29 especies), disminuyendo estas a medida que el bosque fue sometido a algún tipo de intervención. En el BNIR disminuyó a 20 especies y en el BPIR el número de especies observadas fueron solamente 17. En el BTIR el número de especies presentes se redujo considerablemente, encontrándose solo 13 especies. Lo anterior trajo como consecuencia que la frecuencia relativa se distribuyera en un menor número de especies. En el BNI el 55% de la frecuencia relativa se distribuyó en 25 especies de las 29 existentes; en cambio, en el BTIR el 70% de esta frecuencia estuvo representado por sólo 2 especies de las 13 existentes.
El progreso ordenado de una sucesión primaria puede ser interrumpido o hacerse reversible por fuerzas naturales o artificiales (Cronquist, 1974). En el caso del bosque deciduo estudiado, la sucesión primaria, entendiendo por ella el desarrollo de la comunidad vegetal natural hacia la comunidad clímax, fue interrumpida por efecto de la deforestación, posiblemente dando origen a una sucesión secundaria diferente en composición botánica. En las diferentes áreas evaluadas es notorio el efecto de la intervención sobre la diversidad botánica y la frecuencia de las especies, siendo mayor este efecto cuando la ocurrencia del evento fue mas reciente. Cronquist (1974) explicó que las sucesiones en campos abandonados son sucesiones secundarias y que las primeras plantas que invaden el campo por lo general son hierbas anuales. Después de uno o varios años las hierbas anuales dejan paso a las hierbas perennes y estas a su vez dejan el lugar regularmente en unos cuantos años a otras plantas, ya sean árboles, arbustos u otras hierbas.
Con respecto a la densidad de las especies presentes en las diferentes áreas de este bosque, los valores obtenidos se presentan en el Cuadro 6. En el BNI, la especie con mayor densidad fue Simira kluguei (360 plantas/ha) siguiendo a ésta, las especies Guazuma tomentosa, Hecatostemun guazumaefolium y Casearia sp., con valores de 333, 300 y 253 plantas/ha, respectivamente. En el BNIR la especie con mayor densidad fue Apidosperma cuspa, con un valor aproximado de 846 plantas/ha. Según la experiencia local sus frutos son consumidos por los vacunos. En orden descendente de importancia, se encontraron las especies Banisteriopsis cornifolia (487 plantas/ha), Tabebuia serratifolia (160 plantas/ha) y Bourreria cumanensis (133 plantas/ha). En el BPIR, nuevamente la especie Apidosperma cuspa se encontró con mayor densidad, estando presentes alrededor de 693 plantas/ha, seguida por las especies Tabebuia serratifolia (393 plantas/ha), Chloroleucon manguense (273 plantas/ha) y Casearia sp. (247plantas/ha). En el BTIR, las especies con mayor densidad se encuentran asociadas a la deforestación y al posterior abandono del campo, lo cual conlleva a una sucesión secundaria, con predominio de Senna oxyphylla, Acacia macracanta y Acacia articulata (987, 380 y 206 plantas/ha, respectivamente). El BNI fue el área con mayor densidad de plantas por hectárea (2.780 plantas/ha), lo cual estaría indicando una mayor posibilidad de obtener follaje y frutos para la alimentación de rumiantes al estar asociada a un menor grado de intervención. Además, en este bosque la presencia de la familia Leguminoseae es alta, y en su mayoría producen frutos durante la época seca, los cuales son consumidos por los rumiantes, aunque la menor densidad se observó en el BTIR (1.920 plantas/ha), el potencial forrajero de este bosque sigue siendo de interés, dada la presencia de especies con alta producción de frutos, como Acacia macracanta. La producción de frutos para las cuatro especies de leguminosas mas consumidas por los rumiantes de acuerdo a la experiencia local se muestra en el Cuadro 7. Se observaron diferencias en la producción de frutos por especie entre áreas (P<0,05), debido a que la densidad de la especie en cada área fue diferente. Las variaciones en la densidad de especies podrían estar relacionadas con factores que afectan la composición botánica de las áreas, como son la intervención del bosque y la dinámica particular de cada especie. La intervención trae como consecuencia la aparición de especies leguminosas típicas de zonas deforestadas como Acacia macracanta, Senna oxyphylla y Senna atomaria, y la dinámica individual puede estar relacionada con las condiciones climáticas, edáficas o endógenas de la planta, las cuales afectan el desarrollo de la especie en un área determinada. Castro y Magallanes (1993) señalaron que cada especie tiene una dinámica particular que puede verse afectada por las condiciones climáticas o factores endógenos, los cuales influyen sobre la ocurrencia de los estadios reproductivos y por ende su supervivencia a largo plazo.
La mayor producción de frutos se obtuvo para Acacia macracanta en el BTIR (4.500 kg/ha), debido a su mayor densidad poblacional. Esta especie es característica de los campos abandonados en sucesión secundaria. La otra especie importante fue Caesalpinea granadillo en el BPIR, con un valor cercano a los 2.200 kg/ha. La cantidad de frutos producidos en el BTIR fue mayor que en el BNI para las especies estudiadas. No obstante, el bosque no intervenido debería tener una producción de frutos y follaje mayor a la señalada, al considerar todas las especies presentes, ya que de las 29 especies existentes solo fueron evaluadas 4 de ellas. En el Cuadro 8 se presentan los índices de diversidad para las zonas evaluadas. Para el índice de Simpson, el mayor grado de diversidad se observó en el BNI (15,1), con diferencias altamente significativas (P<0,001) con respecto a los del BNIR (5,07), BPIR (6,47) y BTIR (3,15). No se observaron diferencias significativas (P 0,001) entre el BPIR y el BTIR. El BNI presentó una mayor cantidad de especies y un mayor número de individuos. Estas especies son principalmente productoras de frutos y forraje en distintas épocas del año y pueden ser utilizados cuando las deficiencias alimenticias son más críticas, pudiendo constituirse en una fracción importante de la dieta de los animales.
En cuanto al índice de Shannon, la variación en la diversidad de especies entre las diferentes áreas del bosque se comportó de manera similar. El BNI presentó el mayor índice de diversidad (4,01), resultando estadísticamente diferente (P<0,001) al BNIR y al BTIR, los cuales presentaron un índice de diversidad menor (2,80 y 2,0, respectivamente). El BPIR no difirió estadísticamente de los demás grupos. CONCLUSIONES
BIBLIOGRAFÍA
Copyright 2005 - Zootecnia Tropical. The following images related to this document are available:Photo images[zt05010t2.jpg] [zt05010t7.jpg] [zt05010t8.jpg] [zt05010t5.jpg] [zt05010t4.jpg] [zt05010t6.jpg] [zt05010t3.jpg] | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||

{kind=link}