 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Evaluación química de especies no leguminosas con potencial forrajero en el estado Trujillo, Venezuela Chemical evaluation of non leguminous species with fodder potential in Trujillo state, Venezuela Danny E. García1*, María G. Medina2, Carlos Domínguez3, Alfredo Baldizán3, Johny Humbría1 y Luis Cova1
1
Estación Experimental y de Producción Agrícola "Rafael Rangel" Universidad de los Andes, estado Trujillo. Venezuela.
*Correo electrónico: dagamar8@hotmail.com
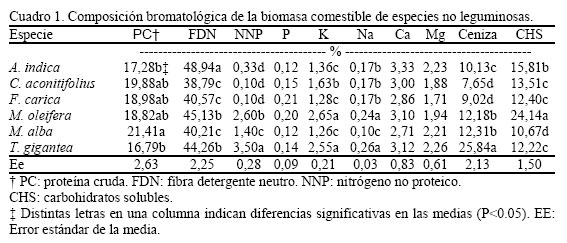
Recibido: 10-02-2006 Code Number: zt06032 RESUMEN Se evaluó la composición fitoquímica de seis especies no leguminosas (Azadirachta indica, Cnidoscolus aconitifolius, Ficus carica, Moringa oleifera, Morus alba y Trichantera gigantea) en el estado Trujillo de Venezuela mediante un diseño totalmente aleatorizado y cinco réplicas. Los contenidos de proteína cruda en todas las plantas fueron elevados y la morera presentó un nivel significativamente superior al resto (21,4%). A. indica sobresalió debido a su fracción fibrosa (48,9%) y T. gigantea exhibió las cantidades más elevadas de nitrógeno no proteico (3,5%). Los niveles de P, Ca y Mg no presentaron variaciones importantes entre las arbóreas y las máximas concentraciones de K y Na se observaron en M. oleifera y T. gigantea (2,65 0,24 y 2,55 0,26%, respectivamente). Estas especies, de forma individual, también presentaron los mayores contenidos de carbohidratos solubles (24,1%) y ceniza (25,8%). Se detectó, indistintamente, la presencia de fenoles, flavonoides, cumarinas, taninos que precipitan proteínas (TPP), taninos condensados (TC), esteroides, terpenos, saponinas, mucílagos, compuestos amargos, cianógenos y alcaloides. Los fenoles; así como los flavonoides, terpenos y esteroides fueron los grupos químicos de mayor distribución. A. indica presentó la máxima cantidad de polifenoles totales (4,21%). A. indica, F. carica y M. oleifera exhibieron cantidades similares de TPP (0,79-0,90%) y TC (1,56-1,77%). A. indica y M. alba sobresalieron por sus concentraciones de alcaloides. Las especies evaluadas presentaron una aceptable composición proximal, poca presencia de metabolitos secundarios protóxicos y bajas concentraciones de posibles factores antinutricionales. Estas especies constituyen buenas alternativas como alimento suplementario en los sistemas de producción en el trópico. Palabras clave: composición química, factores antinutricionales, metabolitos secundarios y especies no leguminosas. Summary The phytochemical compositions of six non leguminous species (Azadirachta indica, Cnidoscolus aconitifolius, Ficus carica, Moringa oleifera, Morus alba, and Trichantera gigantea) were evaluated in Trujillo state, Venezuela, using a totally randomized design and five replicates. The crude protein contents were acceptable and mulberry showed a significantly superior level (21.4%). A. indica stood out due to its fibrous fraction (48.9%) and T. gigantea exhibited the highest quantities of non-protein nitrogen (3.5%). P, Ca and Mg levels did not show important variations and the maximum concentrations of K and Na were observed in M. oleifera and T. gigantea (2.65, 0.24 and 2.55 0.26%, respectively). These species, in individual form, also presented the highest contents of soluble carbohydrates (24.1%) and ash (25.8%). Phenols, flavonoids, coumarins, tannins precipitans of proteins (TPP), condensed tannins (CT), steroids, terpens, saponins, slimes, bitter compounds (B. compounds), cyanogens and alkaloids were detected. Phenols, flavonoides, terpens, and steroids showed a wide distribution. A. indica showed the maximum quantity of total polyphenols (4.21%). A. indica, F. carica, and M. oleifera exhibited similar quantities of TPP (0.79-0.90%) and CT (1.56-1.77%). A. indica y M. alba showed the highest alkaloids concentrations. The evaluated species presented an acceptable proximal composition, low presence of protoxic secondary metabolites and low concentrations of possible antinutritional factors. These species constitute a good alternative as supplementary feed in the production systems in the tropic. Keywords: phytochemical composition, antinutritional factors, secondary metabolites, non leguminous species. INTRODUCCIÓN Recientemente ha surgido un creciente interés en la búsqueda de recursos alimenticios que puedan sustituir parcialmente el uso de concentrados costosos y agroecológicamente distanciados de la realidad ambiental que permitan proveer, de una manera eficiente y económicamente viable, energía, proteína y minerales a los animales herbívoros. Al respecto, las plantas arbóreas y arbustivas tienen un papel preponderante por su elevado valor nutritivo y naturaleza multipropósito. En este sentido existen muchas especies con buenas propiedades forrajeras, entre las cuales se destacan las leguminosas por excelencia (Simón, 1998). No obstante, existen otras leñosas perennes con gran potencial que no han sido empleadas de manera extensiva y su uso ha estado limitado a sistemas de alimentación específicos y aislados. Dentro de ese numeroso grupo se pueden citar al Nacedero o Naranjillo (Trichantera gigantea), la Moringa (Moringa oleifera), el árbol del Nim (Azadirachta indica), las especies de Ficus y Tethonia, la Morera (Morus alba), el Chicasquil, Chaya o Lechosa de jardín (Cnidoscolus sp.) y el Guácimo (Guazuma ulmifolia), principalmente por su gran versatilidad, rápido crecimiento y recuperación después del corte, además de presentar considerables producciones de biomasa en el período seco. Debido a la importancia en el estudio de estas especies para la ganadería tropical se precisa conocer, de manera integrada, los principales indicadores de su composición bromatológica, así como la presencia de posibles compuestos tóxicos y los niveles de metabolitos secundarios presentes en su biomasa. El presente trabajo tuvo como propósito fundamental la evaluación de la composición fitoquímica de seis especies no leguminosas. MATERIALES Y MÉTODOS Características de la zona de muestreo La recolección del material vegetal se realizó en el área forrajera del Módulo de Producción Caprino en la Estación Experimental y de Producción Agrícola "Rafael Rangel", perteneciente a la Universidad de los Andes, en el municipio Pampán, estado Trujillo, Venezuela. La localidad presenta una precipitación promedio anual de 1.200 mm, temperatura media de 28ºC y condiciones de bosque seco tropical. Recolección y preparación de muestras La fracción comestible de hojas y tallos finos (160 días de edad) de las seis especies evaluadas (A. indica, Cnidoscolus aconitifolius, Ficus carica, M. oleifera, M. alba y T. gigantea) fueron colectadas en el mes de febrero del 2005. La totalidad del material se llevó de forma inmediata al laboratorio y fueron secadas a temperatura ambiente, en un local ventilado y oscuro por espacio de doce días. Posteriormente fueron molinadas utilizando una criba de 1 mm y se almacenaron en frascos herméticos hasta la realización del análisis proximal y de metabolitos secundarios. Mediciones analíticas Bromatología A cada muestra se le determinó el contenido de proteína cruda (PC), P, K, Na, Ca, Mg y ceniza según las metodologías propuestas por la AOAC (1990). La fibra detergente neutro (FDN) se determino mediante el protocolo experimental descrito por Van Soest et al. (1991) y los carbohidratos solubles (CHS) y el nitrógeno no proteico (NNP) siguiendo las indicaciones señaladas por Lezcano y González (2000). Tamizaje fitoquímico Se evaluó la presencia de fenoles, flavonoides, cumarinas, quinonas, taninos que precipitan proteína (TPP), taninos condensados (TC), grupos alfa-aminos, cardiotónicos, esteroides, terpenos, saponinas, mucílagos, compuestos amargos (C. amargos), cianógenos y alcaloides. La detección se realizó básicamente según los ensayos cualitativos individuales para cada grupo químico (De Marcano y Hasegawa, 1991) y la metodología clásica de fraccionamiento por polaridad de solvente (Galindo et al., 1989). Cuantificación de metabolitos secundarios La cuantificación de polifenoles totales (FT) se realizó mediante el método de Folin-Ciocalteu, empleando ácido fosfomolíbdico como complejo desarrollador de color (Makkar, 2003). Los TPP se determinaron por el ensayo de la albúmina de suero bovino (Makkar et al. 1988) y los TC mediante el método de nButanol/HCl/Fe3+ y la utilización de acetona (70%) (Porter et al., 1986). La determinación de los alcaloides se baso en la titulación ácida (Sotelo et al., 1996). Diseño experimental y métodos estadísticos Se empleó un diseño totalmente aleatorizado con cinco réplicas. Para el procesamiento de la información se utilizó el paquete estadístico SPSS versión 10.0. A los datos se le realizó un ANOVA usando la dócima de comparación de Student-Newman-Keuls (SNK) y las medias fueron comparadas al 5% de probabilidad. RESULTADOS Y DISCUSIÓN Análisis proximal En el Cuadro 1 se muestran los resultados del análisis bromatológico en las especies evaluadas. Los valores de PC oscilaron entre 16,79 (T. gigantea) y 21,41% (M. alba). A. indica, C. aconitifolius, F. carica y M. oleifera presentaron niveles proteicos intermedios y similares entre sí. En este sentido la cantidad de PC encontrada en las especies es comparable con los niveles clásicos informados en las leguminosas tropicales y en algunos casos superiores a éstas por algunas unidades porcentuales (García, 2003). Teniendo en cuenta los niveles encontrados, algunas especies no leguminosas pueden ser consideradas como alimentos esencialmente proteicos en los suplementos para rumiantes y monogástricos, de igual forma que otras arbóreas clásicamente utilizadas en los sistemas de producción animal tales como Leucaena, Gliricidia y Albizia. Asimismo, los niveles de PC de F. carica, M. alba y T. gigantea coinciden con los reportados en varios ecotipos adaptados a las condiciones de Centroamérica y el Caribe (Pinto et al., 2002; Savón et al., 2005). Por su parte, la FDN fue máxima en A. indica (48,94%), intermedia en M. oleifera y T. gigantea, y significativamente inferiores en las especies de la familia Moraceae (Ficus y Morera). Adicionalmente los niveles encontrados, teniendo en cuenta la edad de la biomasa, son similares a los informados por otros autores en especies del bosque seco tropical de los llanos Venezolanos utilizadas, de manera empírica, como alimento para vacunos (Baldizán, 2004). Los contenidos de NNP también presentaron variaciones importantes entre las especies; A. indica, C. aconitifolius y F. carica exhibieron las menores concentraciones. Por su parte M. alba y M. oleifera, presentaron valores intermedios y diferenciados entre sí y en T. gigantea se observó los mayores índices nitrogenados no derivados de proteínas (3,50%). Los elevados valores de NNP en T. gigantea se encuentran estrechamente relacionados con la singularidad que presentan las especies pertenecientes a la familia Acanthaceae de almacenar considerables concentraciones de sales nitrogenadas inorgánicas en su región aérea (Savón et al., 2005). Los valores de P, Ca y Mg no presentaron diferencias significativas (P<0,05) y los rangos oscilaron entre 0,12-0,21 2,71-3,33 y 1,71-2,25%, respectivamente. Estos niveles de minerales coinciden con los observados en la mayoría de las plantas arbóreas y arbustivas del trópico, aunque dichas concentraciones no cubren los requerimientos de los rumiantes a pastoreo (Colectivo de autores, 2000). Con relación a los elementos K y Na, M. oleifera y T. gigantea presentaron las concentraciones máximas y M. alba el nivel más bajos de Na; el resto de las especies exhibieron cantidades intermedias y diferenciadas entre sí (P<0,05). En cuanto al contenido de ceniza, T. gigantea mostró el mayor aporte; A. indica, M. oleifera y M. alba valores intermedios y C. aconitifolius y F. carica las menores cantidades. No obstante, todos las concentraciones son similares, y en algunos casos superiores, a los informados en las especies forrajeras más comunes en América Central y el Caribe (González y Cáceres, 2002). Por otra parte, las especies presentaron contenidos de CHS muy diferenciados que oscilaron entre 12,22% en T. gigantea y 24,14% en M. oleifera. La elevada variabilidad interespecífica en este indicador energético quizás se deba a la diferenciada capacidad fotosintetizadora de las especies, relacionado con el elemento K (máximo en M. oleifera) el cual es mediador del metabolismo y el transporte de los carbohidratos primarios en las plantas (Pineda, 2004). Detección de metabolitos secundarios De los quince grupos de metabolitos secundarios investigados mediante la utilización de las pruebas cualitativas del tamizaje fitoquímico solo se detectaron en diferentes escalas los fenoles, flavonoides, cumarinas, TPP, TC, esteroides, terpenos, saponinas, mucílagos, C. amargos y alcaloides (Cuadro 2). Estos grupos químicos presentan probada actividad biológica por su posible acción antinutricional en el sistema digestivo de los herbívoros, pero que a su vez pueden ocasionar efectos beneficiosos en dependencia del tipo de compuesto y su concentración en la biomasa (Ramos et al., 1998). La ausencia de grupos alfa-aminos, cardiotónicos y las quinonas es muy positiva, ya que tradicionalmente ocasionan trastornos nutricionales cuando sus niveles son elevados, debido a que los mecanismos de destoxificación no pueden eliminar los metabolitos derivados de estos tóxicos naturales (De Marcano y Hasegawa, 1991). En este sentido los fenoles, flavonoides, esteroides, terpenos y alcaloides estuvieron presentes en la biomasa de todas las plantas evaluadas, mientras que el resto de los metabolitos que fueron detectados solo presentaron una distribución limitada en dependencia de la especie, quizás por presentar funciones muy específicas en las plantas en estudio (García, 2004). La presencia de fenoles y esteroides en todas las especies fue abundante, mientras que los demás metabolitos presentes mostraron una marcada variabilidad en los ensayos cualitativos. Las especies A. indica, M. oleifera y F. carica agruparon la mayor cantidad de metabolitos, mientras que C. aconitifolius, M. alba y T. gigantea tuvieron menor diversidad de estructuras secundarias. Al respecto, es conocido que la poca diversidad de compuestos secundarios en la biomasa comestible de las especies forrajeras constituye, en principio, un buen indicador de palatabilidad (Simón, 1998). En el caso particular de los fenoles, los ensayos individuales se caracterizaron por presentar coloraciones intensas, lo cual denota la amplia diversidad de estructuras hidroxiladas (García et al., 2003). Aunque estos metabolitos no siempre constituyen factores antinutricionales, la presencia de fenoles en plantas de interés agrícola ha sido reportada por muchos autores en pruebas de metabolismo y nutrición. Debido fundamentalmente a que se encuentran ampliamente distribuidos en el reino vegetal formando parte de todas las plantas vasculares como en el caso de Leucaena leucocephala, Calliandra calothyrsus, Acacia cyanophylla, Macroptilium atropurpureum y Lablab purpureus (Makkar y Becker, 1998; Ben Salem et al., 1999). El ensayo cualitativo de flavonoides no mostró una variabilidad marcada entre las especies, resultados que no coinciden con los obtenidos en la comparación de otras plantas mediante rangos de variabilidades en escalas numéricas (Mengcheng et al., 1996). Así mismo detecciones similares se han realizado en Gliricidia sepium, Albizia lebbeck y leguminosas rastreras en los cuales estos compuestos no han causado toxicidad en rumiantes (Martínez et al., 1996). Por otra parte las cumarinas solo fueron detectadas en A. indica y las especies de Ficus y Morera, con un grado de presencia bajo, resultados que difieren específicamente con los obtenidos en las variedades del género Morus (Cubana, Indonesia, Tigreada y Acorazonada) de mayor producción de biomasa en Cuba (García et al., 2003). En este sentido, las cumarinas también han sido detectadas en especies de familias templadas tales como Umbelíferas y Rutáceas y no se conocen efectos beneficiosos en la alimentación de los rumiantes y monogástricos (Berenbaum, 1991). El análisis cualitativo en la detección de triterpenos y esteroides reveló una relativa similitud cualitativa entre las especies. Los esteroides fueron detectados de manera abundante y el ensayo aplicado se caracterizó por una coloración azul verdosa intensa, lo que evidencia la presencia de varios esteroles en el tejido vegetal. La presencia de beta-Sitosterol y Estigmasterol, metabolitos que producen estas tonalidades, han sido clásicamente relacionados con los procesos de activación del crecimiento vegetal y el metabolismo de las hormonas reguladoras en casi la totalidad de las especies vegetales (Valdés y Balbín, 2000). Por tales motivos estas estructuras químicas no deben causar efectos negativos en los animales aún encontrándose en concentraciones importantes; a excepción de los isoprenoides detectados en A. indica ya que quizás, según las pruebas cualitativas, posiblemente tengan estructuras diferentes al resto. La presencia de alcaloides se determinó mediante el empleo de tres reactivos de grupo (Wagner, Hager, Dragendorf) y en todas las especies se detectó su presencia, resultados que apoyan lo planteado en varias investigaciones con relación a que los compuestos alcaloidales se encuentran en la mayoría de los organismos vegetales. Estas estructuras nitrogenadas están diseminadas en las plantas dicotiledóneas, y de forma particular en las leguminosas forrajeras del género Erythrina en las cuales le confieren propiedades deletéreas (Sotelo et al., 1995). Con relación a la presencia de saponinas se observó gran variabilidad entre las especies, con alturas relativas de la espuma entre 5 y 15 mm, equivalentes a contenidos variables de estos metabolitos (Galindo et al., 1989). No obstante, la prueba no resultó concluyente para poder aseverar su presencia específica, ya que el principio del método cualitativo consiste en la disminución de la tensión superficial del medio, por lo que otros compuestos con propiedades estructurales similares en las plantas evaluadas (mucílagos y glucósidos) pudieron crear falsos positivos al respecto. En cuanto a la presencia de mucílagos (mezclas de compuestos complejos) estuvieron presentes solo de manera cuantiosa en las especies de Ficus y Morera, plantas que por excelencia contiene látex y/o gomas. Teniendo en cuenta estos resultados, se debe señalar que ninguna investigación ha informado que, compuestos de esa naturaleza, constituyan factores anticualitativos a excepción de los que presentan propiedades cáusticas e irritantes, los cuales se encuentran ausentes en estos géneros (García, 2003). Por su parte, los compuestos amargos se detectaron cuantiosamente (tres cruces) solo en la biomasa comestible de A. indica. Este resultado afecta desde el punto de vista integral, la calidad de su biomasa para la alimentación de animales monogástricos debido a la elevada presencia de terpenoides de elevado peso molecular que afectan la palatabilidad de la especie (de Marcano y Hasegawa, 1991). Cuantificación de compuestos antinutricionales potenciales Teniendo en cuenta la concentración de FT (Cuadro 3), A. indica mostró un nivel significativamente superior (P<0,05) al resto de las especies (4,21%), lo que a su vez corrobora los resultados del análisis cualitativo. En este sentido, el nivel de fenoles en el Árbol del Nim supera los reportados en M. alba, Inga sp., Difiza sp., Albizia falcataria, Arachi pintoi y G. ulmifolia, entre otras forrajeras importantes (Valerio, 1994; García y Ojeda, 2004). Por su parte F. carica (3,51%) y M. oleifera (3,52%) presentaron niveles intermedios y sin diferencias estadísticas entre sí, que a su vez coinciden con las concentraciones informadas en algunas de las leguminosas de mayor distribución subtropical, tales como Acacia nilotica, Vicia sativa y Quercus incana (Makkar, 2003). Finalmente, las especies que en su conjunto presentaron menor diversidad de metabolitos secundarios (C. aconitifolius, M. alba y T. gigantea) también exhibieron baja fracción de compuestos fenólicos. No obstante, las concentraciones de M. alba y T. gigantea son superiores a las reportadas en otras condiciones de temperatura y suelo (Datta, 2002). De forma global, los contenidos totales de fenoles no superan los límites críticos en los cuales ocasionan daños al buen funcionamiento digestivo de los rumiantes y que ha sido señalado en investigaciones recientes (Makkar, 2003). Con relación a las concentraciones de TPP y TC, no se observaron diferencias significativas entre las especies que presentaron estos metabolitos (A. indica, F. carica y M. oleifera), y los valores oscilaron entre 0,79-0,90 y 1,56-1,77%, respectivamente. A manera de comparación, basado en la utilización de los mismos métodos analíticos estandarizados para plantas forrajeras, las concentraciones máximas de TPP y TC son inferiores a los niveles en los cuales la cantidad de compuestos simples derivados del flavonol-3,4-diol pueden causar trastornos fisiológicos en los rumiantes. No obstante, se requiere profundizar en la determinación de la actividad biológica de los principales tipos de metabolitos polifenólicos para establecer criterios determinantes en estas tres especies. En cuanto al contenido de alcaloides, A. indica y M. alba presentaron los mayores niveles; sin embargo las concentraciones obtenidas son inferiores a las informadas en algunas leguminosas tropicales, las cuales son ávidamente consumidas por la fauna silvestre, el ganado ovino, caprino y bovino (Sotelo et al., 1996; García, 2003). Por tales motivos no deben afectar el consumo voluntario ni repercutir negativamente en la salud animal. La elevada variabilidad estructural de los alcaloides; así como las disímiles propiedades biológicas de estos metabolitos, son factores que se deben estudiar en estas especies para dilucidar su verdadero efecto en la nutrición de los rumiantes. CONCLUSIONES Las especies A. indica, C. aconitifolius, F. carica, M. oleifera, M. alba y T. gigantea constituyen una importante fuente de forraje, dado sus considerables valores de PC, minerales, poca presencia de compuestos protóxicos y relativamente bajas concentraciones de posibles factores antinutritivos. Sin embargo, el follaje de A. indica, dada sus características fitoquímicas, debe ser utilizado para la alimentación de los rumiantes, en los cuales es más factible su uso. RECOMENDACIONES Realizar un uso más intensivo de estas fuentes de alimento y diseñar pruebas de metabolismo en las cuales se determine su potencial de alimentación. No obstante, se requiere profundizar en las características de los principales tipos de metabolitos polifenólicos para establecer criterios determinantes en las especies que los contienen. También se deben realizar evaluaciones de manejo agronómico con la finalidad de determinar la adaptación y productividad de cada especie. LITERATURA CITADA
Copyright 2006 - Zootecnia Tropical |

{kind=link}
{kind=link}
{kind=link}