 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Zootecnia Tropical, Vol. 24, No. 4, 2006, pp. 435-455 Efectos de la ractopamina y lisina sobre la deposición de grasa en cerdos seleccionados magros en la fase de engorde Effect of ractopamine and lysine on lean yield of leaning pigs in the finishing phase Aníbal Pérez, Nestor E. Obispo*, José Palma y Claudio F. Chicco Instituto Nacional de Investigaciones Agrícolas,
Centro Nacional de Investigaciones Agropecuarias.
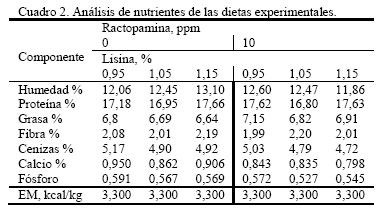
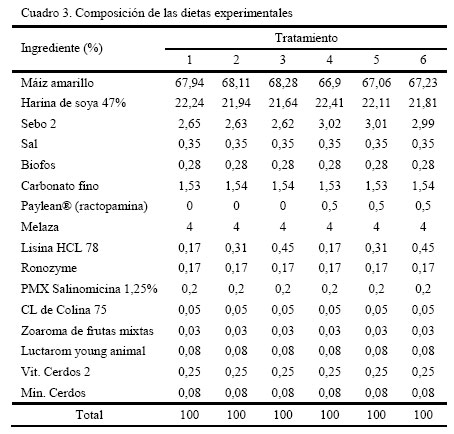
Apartado Postal 4653, Maracay 2101. Aragua. Venezuela. Recibido: 10-11-2005 Code Number: zt06034 RESUMEN De una población de ciento ochenta cerdos, híbridos comerciales mejorados genéticamente para la condición magro, los cuales fueron alimentados con raciones conteniendo dos niveles de ractopamina (RAC: 0 y 10ppm) y tres niveles de lisina (Lis: 0,95, 1,05 y 1,15%) se seleccionó al azar, al momento del sacrificio, una muestra de 60 cerdos (10 por tratamiento, cinco machos y cinco hembras), para evaluar las variables: contenido de magro estimado (TME), profundidad grasa dorsal en última y décima costilla (PG13 y PG10), área del músculo longisimus dorsi, porcentaje de proteína cruda (PC) y de grasa intramuscular (GIM) y relación proteína grasa (P:G). Se observó un efecto entre el nivel de Lys y RAC (P<0,001) sobre PG, siendo más bajo para Lys1,15-RAC10 (1,4 cm) en comparación con Lys0,95 y Lys1,05 con RAC10 (1,73 y 1,68 cm). Así mismo, este tratamiento dio valores más bajo a los niveles de Lys-RAC0 (2,23, 1,74, y 1,72 cm, en el mismo orden de Lys). Se observó un efecto entre Lys y RAC (P<0,05) sobre el PC, siendo el valor más alto a Lys1,15-RAC10, superando por 1,36 unidades porcentuales (UP) a Lys1,15-RAC0. El mismo efecto de interacción (P<0,001) se observó para el GIM con valores más bajos de GIM al nivel de Lys1,15-RAC10 (0,98%) al compararlo con Lys1,15-RAC0 (1,79%) y con 0,95Lys y Lys1,05 con y sin RAC, respectivamente (2,35 2,47 2,25 y 3,22%, respectivamente). Igualmente, se observó efecto entre Lys y RAC (P<0,001) sobre el TME, superando Lys1,15-RAC10 (55,75%) a Lys1,15-RAC0 (52,32%). Se observó un efecto lisina x sexo (P<0,001), sobre el TME, donde las hembras superaron a los machos castrados por 0,41 2,23 y 5,4 UP para Lys0,95, 1,05 y 1,15%, respectivamente. En general, se observó una mejora en la calidad de la canal en cuanto al magro y disminución de la grasa de cobertura dependiente individualmente o en combinación entre el nivel de lisina y ractopamina. Por otro lado, la concordancia de los valores de magro estimados y los de la relación P:G indican que este último podría usarse como un buen indicador de calidad de magro. Palabras clave: cerdo, ractopamina, lisina, porcentaje de magro, grasa, ceba. Summary From a population of one hundred-eighty commercial hybrids pigs, fed diets containing two levels of ractopamine (RAC:0 and 10ppm) and three levels of lisina (Lys:0.95, 1.05 and 1.15%) a sample of 60 animals (10 per treatment, five males and five females) were pulled out at random to evaluate their carcass lean. It was estimated the lean percentage (EL), and the following variables: backfat depth (cm) at the tenth and last rib (DF10 and DF13), the area of longisimus dorsi muscle, protein (CP) and intramuscular fat (IMF) contents (%), and a calculation of the protein:fat ratio. It was observed an effect between of the level of Lys and RAC (P<0.001) on DF, with lower values for Lys1.15 and RAC10 (1.4 cm) as comparing to Lys0.95 and Lys1.05 with RAC10 (1.73 and 1.68 cm). Also, this treatment showed lower values of DF than the same levels of Lys-RAC0 (2.23, 1.74, and 1.72cm, in the same order of Lys). It was observed an interaction between Lys and RAC (P<0.05) on the CP content, with higher values for Lys1.15-RAC10, going above by 1.36 percentage units (PU) the Lys1.15-RAC0 treatment. The same effect (P<0.001) was observed for the IMF, with the smaller values for Lys1.15-RAC10 (0.98%) as compared to Lys1.15-RAC0 (1.79%) and to 0.95Lys and 1.05Lys with and without RAC (2.35 2.47 2.25 and 3.22%, respectively). Also, an effect (P<0.001) between Lys and RAC was observed on the EL, Lys1.15 (55.75%) was better than Lys1.15-RAC0 (52.32%). It was observed an effect (P<0.001) between Lys and Sex on the EL, with better values in the females as compared to the castrated males (0.41 2.23 and 5 PU for Lys0.95, 1.05 and 1.15%, respectively). An improvement on lean and on decreasing fat in the carcasses was related to both Lys and RAC levels (separately or combined). The agreement of EL values and P:G ratios indicate that P:G ratio could be used as good indicator of lean. Keywords: Pig, ractopamine, lysine, dressing percentage, average daily gain. INTRODUCCIÓN La producción de cerdos no sólo ha avanzado en la obtención de líneas genéticas más precoces, con mejores índices de conversión de alimento, sino también y hacia la obtención de cerdos con canales mucho más magras. Este avance ha sido motivado primeramente a la necesidad de incrementar los rendimientos obtenidos en el desposte de las canales, de manera de proporcionar una mayor cantidad de carne con la consecuente mejora en la rentabilidad, y a una mayor demanda de este tipo de carnes en base a exigencias relacionadas con la salud de los consumidores. Adicionalmente a la mejora genética, se han desarrollado aditivos no nutricionales, los cuales son capaces de inducir adicionales respuestas en la calidad de las canales, como por ejemplo el compuesto ractopamina (RAC), un agonista β-adrenérgico de la familia de las fenoletanolaminas, que actúa sobre los receptores β-adrenérgicos de las células adiposas y del músculo esquelético, promoviendo la lipólisis, con el consecuente incremento del magro en la canal (Smith y Paulson, 1994; Spurlock et al., 1994; Crome et al., 1996). El empleo de RAC ha permitido experimentar resultados variables sobre el cerdo al sobre la respuesta productiva, ganancia diaria de peso (Dunshea et al., 1993; Williams et al., 1994), rendimiento en canal (Williams et al., 1994; Crome et al., 1996; Pérez et al., 2005) y pérdidas por goteo (Pérez et al., 2005). Por otro lado, al hablar de los requerimientos de aminoácidos en la alimentación de los cerdos, hay que tomar en cuenta que están basados en suplir en primera instancia los requerimientos de lisina, el cual ha sido considerado como el principal aminoácido limitante en la alimentación de esta especie (Batterham et al., 1990; Bikker et al., 1994). Sin embargo, otras investigaciones consideran que los aminoácidos azufrados (AAS) metionina y cisteína también son limitantes, por lo que entonces, será importante mantener una óptima proporción AAS:lisina (Knowles et al., 1998). Se ha considerado que para minimizar la deposición de grasa y garantizar un buen crecimiento y desarrollo muscular, esta proporción debería estar cercana al 0,67%. Aunque parece no ser así, en aquellos animales alimentados a base de maíz, harinas de soya o sorgo, los cuales dependerán más del nivel de lisina suministrado (Knowles et al., 1998). Por consiguiente, la distribución del resto de los aminoácidos en la proteína va a depender de la cantidad de lisina requerida por el cerdo en su respectivo estado fisiológico constituyéndose de esta manera en lo que se ha llamado la proteína ideal. Concomitante al efecto de la selección genética para la obtención de animales mucho más magros, se ha propiciado un aumento de los requerimientos de lisina del cerdo (Friesen et al., 1994). Por lo tanto, los requerimientos de lisina para los cerdos durante el engorde dependerán del tipo de dieta y del criterio de respuesta, en el intento de obtener una óptima respuesta en ganancia de peso, conversión de alimento, características de la canal y tasa de retención de proteína en el músculo para lo cual es importante establecer un nivel adecuado nivel de este aminoácido en la dieta (NRC, 1998) Aparte de los factores genéticos, los requerimientos de aminoácidos en los cerdos se encuentran influenciado por otros, tales como sexo, estado fisiológico, concentración de energía de la dieta, biodisponibilidad de estos aminoácidos y la frecuencia de alimentación (Hahn et al., 1995). Así, se han evaluado los efectos de la lisina sobre las características de la canal del cerdo, sobre el rendimiento de la canal al beneficio, profundidad de la grasa dorsal en la última y antepenúltima costilla, área del músculo longisimus dorsi y porcentaje de tejido magro, encontrándose resultados variables de acuerdo al nivel de incorporación dado, capacidad genética y el sexo (Hansen y Lewis. 1993; Hahn et al., 1995). Igualmente, se ha observado incrementos en la grasa intramuscular en el lomo del cerdo con dietas deficientes en lisina (Cisneros et al., 1996; Witte et al., 2000). Por otro lado, algunos estudios (Schinckel et al., 2003) señalan que el nivel de lisina suministrado en la dieta afecta la magnitud de la respuesta de RAC sobre las características de la canal. El objetivo de esta investigación fue evaluar el efecto de la ractopamina a tres diferentes niveles de lisina sobre la modificación del tejido magro de cerdos en la fase de engorde. MATERIALES Y MÉTODOS El experimento se realizó en la granja "Los 333" del grupo "La Caridad C.A." ubicada en las afueras de la localidad de Parapara de Ortiz, municipio Roscio del estado Guarico, Venezuela. De una población de 3.500 cerdos mejorados genéticamente de la línea Pig Improvement Company (PIC), híbrido comercial proveniente del cruce de las razas Landrace, Duroc, Large White, Hampshire, Berkshire, Pietrain y Meishan, se seleccionaron 180 animales con una edad y peso inicial de 156 días y 93 ± 10 kg, respectivamente. Los cerdos fueron alojados a razón de 30 animales por corral, asignados al azar, considerándose la talla para minimizar los efectos de dominancia dentro del corral. En base a los registros de humedad y temperatura de la unidad de producción se consideró que, a lo largo y ancho de los galpones, los diferentes corrales fueron considerados homogéneos en la condición microambiental. Cada cerdo fue identificado en la oreja derecha de manera de facilitar la recolección de los datos. Los animales fueron asignados a 12 tratamientos en base a un diseño completamente aleatorizado con arreglo tipo factorial 3x2, tres niveles de inclusión del aminoácido lisina (Lis: 0,95, 1,05 y 1,15%) y dos niveles del aditivo ractopamina (RAC: 0 y 10 ppm). Sin embargo, hay que acotar que debido a requerimientos de la granja, cada corral debió conformase con una proporción igual de machos castrados y hembras, por lo que el factor sexo también fue considerado. Los diferentes tratamientos se muestran en el Cuadro 1. Los cerdos tuvieron acceso libre al aliento y al agua de bebida. Las dietas fueron formuladas para ser isoproteicas (17,5% PC) e isoenergéticas (3.300 kcal EM/kg) (Cuadro 2) y su composición se muestra en el Cuadro 3. Las determinaciones para esta investigación se realizaron sobre una muestra de diez canales seleccionada al azar a nivel de matadero de cada una de los tratamientos anteriormente descritos (cinco machos castrados y cinco hembras). El día 27, último día del ensayo, los animales fueron sometidos a un proceso de ayuna por 16 horas, a de manera de garantizar el máximo de desocupación de las vísceras intestinales. Cada grupo fue trasportado al matadero, efectuándose el sacrificio por aturdimiento a través de choques eléctricos, degollado, escalado y desviscerado, preservándose en las canales la piel, cabeza y extremidades. Para las determinaciones de laboratorio, diez canales por tratamiento fueron seleccionadas al azar (5 de cada sexo), a las cuales les fue tomada una sección del lomo de la canal derecha entre la 10ma y la 13ra costillas (Dunshea et al., 1993). Las muestras fueron empaquetadas en bolsas plásticas y refrigeradas por 72 horas a 4°C para luego realizar las mediciones de las variables de modificación del magro. La profundidad de la grasa dorsal a nivel de la décima (PG10) y décimo tercera (PG13) costilla (cm), se realizó utilizando un Vernier, midiéndose perpendicularmente desde el mismo punto, en ambos extremos, a ¾ de distancia de la línea media de las vértebras. Esta medición correspondió al espesor de la grasa subcutánea y a la correspondiente de piel y el tocino. Posteriormente, se procedió a copiar (calcado) en un papel cebolla la figura del músculo longisimus dorsi, del lado correspondiente a la 10ma costilla, luego de recortada la misma fue medida en un lector de área foliar (LI-COR®) para determinar el área del músculo en cm2 (AMLD). Para la determinación del porcentaje de tejido magro, se procedió a aplicar la ecuación propuesta por el Consejo Nacional de Productores de Cerdos de los Estados Unidos (National Pork Producers Council, 1994), la cual considera las variables peso frío de la canal, corregida por el factor 0,985 (corrección del peso frío de la canal peso frío a caliente), el área del músculo longisimus dorsi en la 10ma costilla y la PG10: Los análisis químicos se realizaron en el Laboratorio de Nutrición Animal del Instituto Nacional de Investigaciones Agrícolas en Maracay, estado Aragua, determinándose la concentración de proteína cruda en el músculo, a través del método de Kjeldahl, y la grasa intramuscular por extracción en Soxtec (AOAC, 1989). Con estos valores se calculó relación proteína:grasa. Para la evaluación estadística se utilizó el siguiente modelo lineal: Yijk = μ + αi + βj + (αβ)ij +(a ρ)ik + (b ρ)jk + ρk + εijk
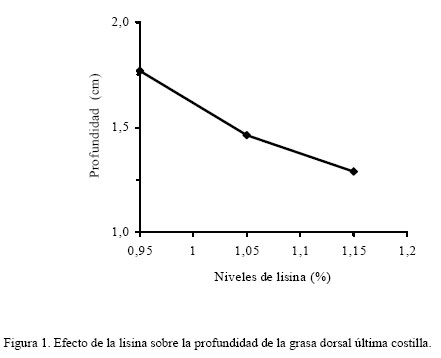
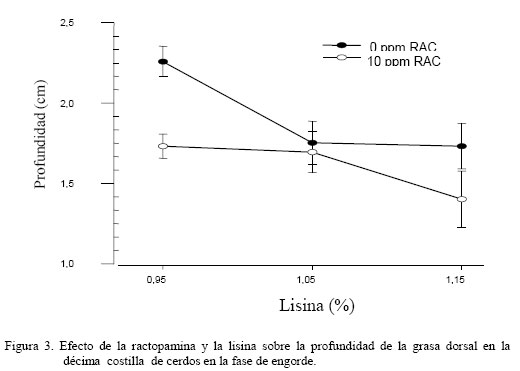
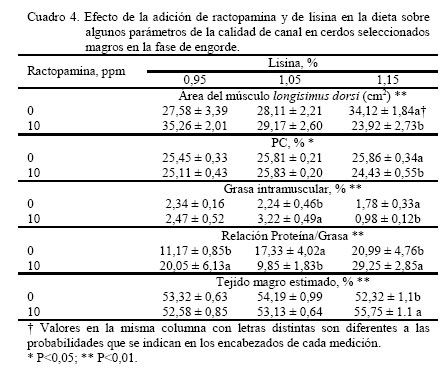
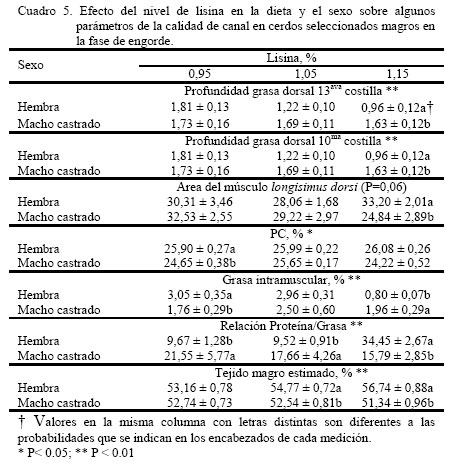
donde: Yijk = Respuesta productiva en el i–ésimo nivel de lisina, j–ésimo nivel de RAC y k–ésimo sexo. μ = Media Teórica de la población. αi = Efecto del i-ésimo nivel de lisina ( i = 1,...3) βj = Efecto del j-ésimo nivel de RAC ( j = 1,.2) ρk = Efecto del k-ésimo sexo (k = 1,..2) (αβ)ij = Efecto de la interacción de primer orden del i-ésimo nivel de lisina y el j-ésimo nivel de RAC. (αρ)ik = Efecto de la interacción de primer orden del i-ésimo nivel de lisina el k-ésimo sexo. (βρ)jk = Efecto de la interacción de primer orden del i-ésimo nivel de RAC y el i-ésimo sexo. εijk = Error experimental del i–ésimo nivel de lisina, el j-ésimo nivel de RAC y la k-ésimo sexo. El análisis de varianza fue realizado empleando el programa estadístico SAS (1985) bajo el procedimiento GLM, las medias fueron ajustadas por mínimos cuadrados y las medias se compararon a través de la prueba de rango múltiple de la mínima diferencia significativa (Steel y Torrie, 1980). RESULTADOS Y DISCUSIÓN El análisis estadístico no indico interacción triple significativa entre sexo, RAC y el nivel de lisina de la dieta. Independiente del nivel de RAC, al nivel 1,15% de lisina, se observó (Figura 1) que PG13 fue menor (P<0,001) al compararla con los niveles de 0,95 y 1,05% (1,29 vs. 1,77 y 1,46 cm, respectivamente). Con el incremento del nivel de lisina en la dieta, la PG13 tiende a disminuir, posiblemente motivado a un incremento en la tasa de retención del nitrógeno en el músculo (Hansen y Lewis, 1993; Hahn et al., 1995; Chen et al., 1999). Así mismo, la inclusión de RAC disminuyó (P<0,001) la PG13 en comparación con el tratamiento control (1,33 vs. 1,69) (Figura 2). Estos resultados son coincidentes con los reportados por Dunshea et al. (1993); Crome et al. (1996) y Schinckel et al. (2003). La PG10 en relación al nivel de lisina no fue independiente del nivel de RAC (P<0,05), siendo este valor más bajos al nivel 1,15% de lisina y 10 ppm de RAC (1,4 cm) (Figura 3), observándose un efecto sinergístico hacia la disminución de la grasa de cobertura de entre lisina y RAC, lo que es coincidente con lo observado por Dunshea et al. (1993), Crome et al. (1996) y Schinckel et al. (2003). Al comparar el AMLD (Cuadro 4), se observó al nivel más bajo de lisina sin RAC el valor más alto (P<0,001) con respecto de su par con RAC (35,27 vs. 27,59 cm2). Esta respuesta se invirtió al incrementarse el nivel de lisina, (34,12 vs. 23,93 cm2). En términos generales, se observó que hay una tendencia a disminuir el AMLD en los tratamientos sin RAC al incrementarse el nivel de lisina, y hacia el aumento al añadirse la RAC a mayores niveles de lisina (Cuadro 4). Esta respuesta resultó diferente a los resultados aportados por Schinckel et al. (2003), quienes observaron una tendencia de aumentó del AMLD a medida que se incrementaba el nivel de lisina (0 0,82 y 1,08%) en combinación con 20 ppm de RAC. Sin embargo, es importante considerar que los efectos de la RAC están influenciados por la sensibilidad de tejido adiposo y la baja regulación del β-adrenoreceptor (Liu et al., 1994). Igualmente, la concentración de proteína cruda (PC) fue dependiente del nivel de lisina y RAC en la dieta (P<0,05), donde la inclusión de RAC indujo a una mayor concentración de PC en el músculo, la cual incrementó al aumentar el nivel de lisina suministrado en 25,45 25,81 y 25,86% a 0,95 1,05 y 1,15%, respectivamente, contra los mismos niveles de lisina sin RAC 25,11 25,83 y 24,44% (Cuadro 4). Lawrie (1998) reporto un contenido de PC en el músculo de 22,5%. La RAC induce el incremento del gen transcriptor a -actina y al incremento de ácido ribonucleico mensajero (ARNm) y probablemente otras proteínas miofibrilares, las cuales incrementan la síntesis proteica y disminuyen la degradación proteica (Grant et al., 1995). Es importante señalar que la retención de N es un proceso energético dependiente, en donde el potencial de retención nitrogenado se alcanza a un determinado nivel de ingestión energética (Noblet y Henry, 1991) La concentración de grasa intramuscular (GIM) fue mucho más variable con respecto a la RAC y el nivel de lisina (Cuadro 4), no observándose diferencias al nivel más bajo de lisina, para ambos casos (con y sin RAC); sin embargo, se observó un efecto incremental (P<0,001) al elevarse el nivel de lisina a 1,05% (3,22%), siendo diferente (P<0,01) al mismo nivel sin RAC (2,24%). La GIM disminuyó significativamente al incrementarse el nivel de lisina a 1,15%, valor que resulta igualmente mucho menor al compararlo con el tratamiento sin RAC (0,983 vs. 1,789%). Es muy probable que este tipo de respuesta en los cerdos de este experimento haya sido motivada a la capacidad genética para el magro en este tipo de cerdo. El marmóreo y las características y sensoriales de las canales (dulzura, jugosidad y sabor agradable) han sido asociadas con esta GIM (Castell et al., 1994). Es decir altos valores son algunas veces predilección de algunos consumidores. En esta investigación, los valores obtenidos para esta variable tendieron hacia la baja y se corresponden con la tendencia observada en otras investigaciones, en las cuales se han empleado dietas deficientes en lisina para incrementar el marmóreo (Cisneros et al., 1996; Witte et al., 2000). La relación P:G (Cuadro 4) resultó dependiente de la relación lisina-ractopamina (P<0,001). La relación P:G del tratamiento que recibió RAC al nivel más bajo de lisina resultó superior (P<0,01) en comparación con el par sin RAC (20,05 vs. 11,18). Cuando se incrementó la lisina a 1,05% en el tratamiento con RAC, la relación P:G disminuyó sustancialmente con respecto al nivel más bajo de lisina y con respecto su par sin RAC (9,85 vs. 17,33). A un nivel mayor de lisina (1,15%) con RAC la relación P:G se incrementó (P<0,01) en comparación con respecto a todos los tratamientos (29,25). Sin embargo, el valor de la relación P:G al nivel 1,05% de lisina en el tratamiento con RAC (Cuadro 4), se obtuvo una respuesta inferior a la de su par sin RAC, la cual no es posible explicar al momento. En términos generales, se podría decir que a niveles crecientes de lisina y 10 ppm de RAC se favoreció la acumulación de proteína en el músculo. Como era de esperarse, el TME (Cuadro 4) resultó igualmente dependiente del nivel de lisina y RAC (P<0,01). Al nivel de 1,15% de lisina el tratamiento con RAC mostró un valor más alto (55,75%) en comparación con su par sin RAC (52,32%). A niveles inferiores de lisina, no se apreciaron diferencias por efecto de la adición o no de RAC. Esta respuesta hace evidente que el efecto de la RAC esta influenciada por el nivel de lisina (Schinckel et al., 2003). Igualmente, con esta estimación se pone en evidencia la similitud entre la respuesta de P:G para indicar la magrosidad de la canal. Cuando se comparo el nivel de lisina en la dieta con el sexo se observó una interacción (P<0,001) para la medición de PG13, siendo mucho menor para las hembras (1,22 y 0,95 cm) que para los machos castrados (1,69 y 1,63 cm) a los niveles 1,05 y 1,15% de lisina, respectivamente (Cuadro 5). Los resultados obtenidos en este experimento fueron inferiores a los reportados por Hansen y Lewis (1993) y Hahn et al. (1995); sin embargo, se destaca en ambas experiencias que las hembras tienden a tener un PG13 menor que el de los machos castrados, la cual tiende a disminuir al incrementar la concentración de lisina en la dieta. Análogamente, el mismo efecto se observó para PG10 (Cuadro 5). En general, se observa un efecto uniforme en la medición a lo largo de la sección del lomo evaluada (entre la 10ma y 13ra costilla). Los resultados para PG10 fueron inferiores a los obtenidos por Hansen y Lewis (1993). Los resultados de la AMLD con respecto al nivel de lisina fueron igualmente dependientes del sexo (P=0,06). Al nivel 1,15% de lisina, las hembras mostraron una mayor AMLD con respecto a los machos castrados, superándolos por 8,37 cm2 (Cuadro 5). A los niveles más bajos de lisina no se detectaron diferencias entre los sexos en estudio. Hansen y Lewis (1993), Hahn et al. (1995) y Chen et al. (1999) observaron incrementos del AM por parte de las hembras sobre los machos castrados al incrementar la concentración de lisina en la dieta. El contenido de proteína con respecto al nivel de lisina no fue independiente del sexo (P<0,05), resultando más alto en las hembras con respecto a los machos castrados superándolos 1,24 y 1,85 unidades porcentuales a los niveles 0,95 y 1,15%, respectivamente. Este valor fue similar entre ambos sexos al nivel 1,05% (Cuadro 5). Estos resultados, en líneas generales, fueron superiores al presentado por Lawrie (1998) de 22,5% PC. Parece existir una mayor eficiencia por parte de las hembras en retener nitrógeno con respecto a los machos castrados (Noblet y Henry, 1991). Los resultados de la GIM fueron variables pero dependientes del nivel de lisina y sexo (P<0,001) (Cuadro 5), en donde los machos castrados presentaron una disminución en la GIM con respecto a las hembras al nivel 0,95% (1,75 vs. 3,06% para machos castrados y hembras, respectivamente). Sin embargo, al nivel 1,05% no se observaron diferencias entre machos castrados (2,50%) y hembras (2,97%). Al máximo nivel de lisina las hembras presentaron una disminución en GIM con respecto de los machos castrados (0,80 vs. 1,97%). Estos resultados fueron inferiores a los reportados por Cisneros et al. (1996) y Witte et al. (2000), empleando niveles de lisina más bajos que los de esta investigación. La menor GIM pudiera estar propiciada por la mayor cantidad de lisina ingerida y a la mayor capacidad genética de los cerdos empleados en este ensayo para retener el nitrógeno ingerido. La relación P:G fue igualmente dependiente del nivel de lisina y el sexo (P<0,001), siendo esta relación más alta en los machos castrados con respecto a las hembras a los niveles de 0,95 y 1,05% (21,55 vs. 9,67 y 17,66 vs. 9,52 para machos castrados y hembras, respectivamente). Sin embargo, al nivel 1,15% de lisina las hembras superaron (P<0,001) a los machos castrados (34,46 vs. 15,79) (Cuadro 5). Esta relación se corresponde con los resultados anteriormente descritos de los valores de acumulación de proteína en relación al nivel de lisina y sexo, considerándose que el nivel energético de las dietas fue suficiente para fomentar una mejor retención de nitrógeno (Medel y Fuentetaja, 2001) Se observó un efecto entre el nivel de lisina y sexo para TME (P<0,01), siendo más alto en el caso de las hembras por 2,13 y 5,4 unidades porcentuales para los niveles 1,05 y 1,15%, no observándose diferencias al nivel 0,95% de lisina (Cuadro 5). Estos valores de TME fueron superiores a los reportados por Hansen y Lewis (1993), quienes observaron una tendencia de incrementar esta estimación a medida que se incrementaba el nivel de lisina en la dieta. Según Lopes et al. (2001), esto pudiera ser debido a la carencia en los machos castrados del nivel de andrógenos necesarios para inducir a las receptores androgénicos del músculo esquelético. Para la PG10 no se observó efecto entre el nivel de RAC y el sexo; sin embargo, hubo un efecto (P<0,001) de la RAC sobre la PG10 con una disminución este valor en 0,30 cm en los animales tratados (Cuadro 6). Esto coincide con las observaciones de Dunshea et al. (1993), Crome et al. (1996) y Schinckel et al. (2003). La inclusión de RAC influyó (P<0,05) en la respuesta de PC, siendo más alto en los animales tratados con la RAC superando a los animales sin RAC por 0,59 unidades porcentuales (Cuadro 6) El AMLD presentó un efecto de RAC x sexo (P<0,05), siendo este valor más alto en las hembras sin RAC en comparación con los machos castrados sin RAC, superándolos 5,28 cm2. Sin embargo, la inclusión de 10 ppm de RAC mejoró este valor en los machos castrados (30,92 cm2), sin ser diferente a los observados en las hembras al mismo nivel de RAC (28,96 cm2). En las hembras hubo una tendencia contraria a la observada en los machos castrados (Figura 4). Igualmente se observó un efecto (P<0,001) del sexo sobre la PC, en donde las hembras mostraron un incremento de 1,14 unidades porcentuales con respecto a los machos castrados (Cuadro 7). Estos resultados son coincidentes con los reportados por Weatherup et al. (1998). Es posible que este efecto se encuentre vinculado a la baja acción androgénica en los machos castrados, lo que conlleva a una disminución de la síntesis proteica (Lopes et al., 2001). No se observó efecto significativo entre el nivel de RAC y el sexo sobre las variables GIM y P:G; así mismo, no se apreciaron efectos principales para cada una de ellas. De igual manera, los resultados de la concentración de tejido magro estimado no presentaron interacción entre el nivel de RAC y el sexo del animal; sin embargo, se apreció un efecto del sexo sobre esta variable, en donde las hembras mostraron una más alta TME (P<0,001), superando a los machos castrados por 2,68 unidades porcentuales (Cuadro 7), coincidiendo en los resultados aportados por Weatherup et al. (1998) y Medel y Fuentetaja (2001). Se observó un efecto del sexo (P<0,001) sobre la PG13, en donde las hembras presentaron una disminución 0,36 cm con respecto a los machos castrados (Cuadro 7). Este efecto del sexo coincide con lo observado por Weatherup et al. (1998) y Medel y Fuentetaja (2001). Igualmente, se observó un efecto (P<0,001) del sexo sobre la PG10, observándose en las hembras un valor de 0,42 cm menor de PG10 al compararlas con los machos castrados (Cuadro 7). CONCLUSIONES
LITERATURA CITADA
Copyright 2006 - Zootecnia Tropical |

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}