 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Zootecnia Tropical, Vol. 25, No. 1, 2007, pp. 19-28 Efecto del toro sobre el comportamiento estral de vacas Chinampas (Bos taurus) en una región tropical seca Effect of bull on estrual behavior of Chinampo (Bos taurus) cows in a dry tropical region José Luís Espinoza Villavicencio1*, Rigoberto López Amador1, Alejandro Palacios Espinosa1, Ricardo Ortega Pérez1, Narciso Ávila Serrano2 y Bernardo Murillo Amador3 1 Universidad Autónoma
de Baja California Sur, Carretera
al Sur km. 5.5, CP 23000, La
Paz, BCS, México.
*Correo electrónico: jlvilla@uabcs.mx Recibido: 09/10/2006 Code Number: zt07003 RESUMEN Para conocer el comportamiento estral del bovino Chinampo se utilizaron 24 vacas ciclando. Un grupo (GI) estuvo con un toro protegido con un mandil de tela para evitar la cópula, mientras que el otro (G2) permaneció aislado del macho. Se contabilizaron intentos de monta, montas realizadas, montas recibidas no aceptadas, montas recibidas aceptadas (MA), lamidos, topeteos, acompañamientos y olfateos. Se analizaron sus ocurrencias acumuladas dentro de las 30, 60 y 90 h posteriores al inicio de una observación constante, que inició a partir de las 30 h de concluido un protocolo de sincronización de estros. Dentro de las 30-60 h de observación constante hubo más eventos en G1. Hubo más interacciones secundarias del estro dentro de las 30-60 h en G1. Hasta las 30 h se registraron más MA en G1 y entre 60-90 h se constataron más MA en G2. El 60% de las vacas de G1 y el 18% de G2 iniciaron estro entre las 6:01-12:00 h. En G2, la mayoría de los estros iniciaron durante la noche. El 60% de las MA en G1 ocurrieron entre las 6:01-12:00 h, proporción superior a la observada en G2. La proporción de vacas con duración del estro menor de 8 h fue mayor en G1. Se concluye que en presencia del macho, las vacas Chinampas concentraron su conducta estral en un periodo más corto y aumentó la proporción de estros con duración menor de 8 h. El macho afectó la hora de inicio del estro y concentró las MA durante la mañana. Palabras clave: Efecto macho; Comportamiento estral; Vacas Chinampas. ABSTRACT In order to know the estrual behavior of the Chinampo cows, 24 cycling cows were used. A group (G1) was with a teaser bull, while the other one (G2) remained isolated from the male. Attempts to mount, made mounts, received but not accepted mounts, received and accepted mounts (AM), licking, butting, following, and smelling were recorded. Their accumulated occurrences were analyzed within the 30, 60 and 90 h after the beginning of a constant observation, since the 30 h after a protocol of estrous synchronization was finalized. Within the 30-60 h there were more events in G1. All of the events in both groups happened within 90 h. There were more secondary estrous interactions within the 30-60 h in G1. Until the 30 h, more AM in G1 was registered and between 60-90 h more AM in G2 were stated. Sixty percent of the cows of G1 and 18% of G2 initiated the estrus between 6:01-12:00 h. In G2, most of the estrus happened during the night. Sixty percent of the AM in G1 happened between 6:01-12:00 h, superior proportion to the one observed in G2. The proportion of cows with estrus duration minor of 8 h was greater in G1. We concluded that in male presence, the estrual behavior of Chinampo cows was concentrated in a shorter period and increased the proportion of estrus with minor duration of 8 h. The male affected the time of the estrus beginning and concentrated the AM during the morning. Keywords: Male effect; Estrual behavior; Chinampo cows INTRODUCCIÓN En la República Mexicana existe una gran diversidad genética de bovinos, contándose con diferentes grupos raciales adaptados a las diversas regiones agroecológicas; sin embargo, muchas de estas razas aún no están caracterizadas. El ganado autóctono posee rasgos valiosos como su resistencia a enfermedades endémicas, longevidad y adaptación a situaciones difíciles y a alimentos de baja calidad, características todas ellas deseables para una ganadería sostenible con bajos insumos (Espinoza, 2004). El ganado bovino criollo o Chinampo de la península de Baja California en México no es la excepción; tiene su origen en los bovinos que fueron introducidos por los Jesuitas en 1697 y a través de los siglos se ha generado un tipo de animal pequeño (las vacas adultas pesan alrededor de 250 kg) y se han adaptado al ecosistema (Martínez-Balboa, 1980). Este ganado solamente ha sido tipificado en ciertas características morfométricas (Espinoza et al., 2002) y no se conocen muchas de sus particularidades fisiológicas como la reproducción, a pesar que una comprensión del ciclo estral y el conocimiento de las señales de estro son indispensables para mejorar la eficacia reproductiva (Allrich y Khutson, 1997). El reconocimiento del comportamiento de la vaca durante el estro permite el diseño de programas biotecnológicos como la inseminación artificial, la transferencia de embriones y la monta controlada. El clima, la raza, el sistema de manejo y el grado de interacción social del hato influyen sobre la conducta, lo que ayuda o dificulta la determinación del ciclo (Mai et al., 2002). Es necesario conocer y precisar las diferentes actitudes de las vacas durante el estro ya que se han destacado diferencias importantes entre razas, sobre todo entre ganado Bos taurus y Bos índicus, incluso dentro del mismo ganado europeo dependiendo de determinados factores medio ambientales. Esta información se hace indispensable cuando se utiliza la inseminación artificial como herramienta para el mejoramiento genético. En dichas circunstancias, algunas veces es necesario implementar ciertas medidas como ayudas para la detección de estros. Dentro de esas medidas está el uso de toros marcadores. Sin embargo, no se sabe si en el caso del ganado bovino Chinampo, la presencia del macho en un grupo de vacas con celo sincronizado pudiera alterar su comportamiento estral. Por las razones expuestas anteriormente, el objetivo de este estudio fue determinar las manifestaciones características del comportamiento estral en vacas Chinampas y evaluar el efecto de la presencia del toro sobre su conducta, inmediatamente antes y durante el estro. MATERIALES Y MÉTODOS Localización del área de estudio La presente investigación se realizó en la posta zootécnica de la Universidad Autónoma de Baja California Sur, en La Paz, B.C.S., México. Se encuentra ubicada en una región de trópico seco en las coordenadas geográficas 26º 06´01" N y 110º 0´0" O, a 33 msnm. El clima predominante en la zona, según la clasificación de Köppen, modificado por García (1973) es BW (H) HW (X), siendo este clima seco y calido con lluvias en verano, invierno y escasas todo el año, con una precipitación pluvial media anual de 195.4 mm y una temperatura media anual de 28.7ºC (Dgtenal, 1980). Diseño del estudio Se utilizaron 24 vacas Chinampas de primer parto (40 meses de edad, aproximadamente), sin cría, con un peso promedio de 250 ± 9 kg, las cuales se mantuvieron en corral, con una alimentación consistente en heno de alfalfa (Medicago sativa) conteniendo un 17% de PC y agua a libre acceso. Las vacas, cuyas crías habían sido destetadas a los 7 meses de edad y permanecían vacías, fueron observadas para la detección de estros durante tres ciclos estrales consecutivos con el fin de asegurar que estuvieran presentando un celo cada 18 a 21 d. Antes del experimento, todas las vacas permanecieron separadas del macho en un corral alejado unos 50 m del mismo y manteniendo solamente contacto visual con este. Todos las hembras recibieron una inyección intramuscular de 25 mg de Dinoprost (Lutalyze; Pharmacia & Upjohn) en la mañana (7:00 h) del primer día del tratamiento, repitiéndose la dosis a la misma hora 11 d después de la primera aplicación. Inmediatamente después se formaron dos grupos de vacas aleatoriamente y se mantuvieron en dos corrales separados (10 m de distancia uno del otro), entre los cuales los animales podían verse y escucharse. A partir de ese momento, el grupo 1 (GI; n = 12) se mantuvo junto con un toro protegido en su abdomen por un mandil de tela para evitar la cópula. El grupo 2 (G2; n = 12) permaneció aislado del macho. El toro utilizado (48 meses de edad) fue un híbrido de 430 kg, con experiencia sexual y producto de la cruza de Chinampo con raza(s) Bos taurus para carne, no definida. La observación de los animales se hizo cada 6 h (12:00, 18:00, 24:00 y 6:00 h) después de la segunda inyección de Dinoprost, hasta las 30 h. A partir de entonces, la observación se llevó a cabo de manera permanente durante 90 h adicionales, completando un periodo de observación de 120 h. Todos los animales contaban con un arete de identificación con su número respectivo y antes de iniciar la observación se marcó con pintura el número correspondiente a cada vaca en ambos costados. Durante la noche se utilizó luz artificial tenue para facilitar la observación y el observador se mantuvo alejado para no interferir con el comportamiento de los animales. Las variables evaluadas fueron: intentos de monta (IM) de un animal a otro sin llegar a completarla; montas realizadas (MR) o concluidas sobre animales en celo o no, montas recibidas no aceptadas (MRNA) en las cuales un animal era montado por otro retirándose inmediatamente y rechazando la monta; montas recibidas aceptadas (MA), en este caso la vaca permaneció inmóvil al ser montada por otra; y otras manifestaciones asociadas al estro como, lamiendo (LAM), topeteo (TOPE), acompañamientos (ACO) y olfateo (OLF). En G1 se contabilizaron tanto los eventos realizados por el toro, como aquellos efectuados por las vacas del grupo. Para fines de interpretación se consideró que una vaca se encontraba en celo mientras aceptara la monta (interacción primaria) de sus compañeras o del toro, en su caso (Villagómez et al., 2000). El resto de las interacciones mencionadas con anterioridad se consideraron como signos secundarios o asociados del estro. Análisis estadístico La frecuencia acumulada de cada variable dentro de periodos de 30 h se comparó entre tratamientos (G1 y G2) a través de una prueba de Z para comparación de proporciones a las 30, 60 y 90 h, después de iniciada la observación permanente de los animales. El intervalo entre montas aceptadas, total de interacciones secundarias (IM, MR, MRNA, LAM, TOPE, ACOM y OLF), MA por tratamiento y la duración del estro (DE), considerando a este último como el intervalo en horas entre la primera y la última monta aceptada, se compararon entre grupos mediante el siguiente modelo lineal general: donde:
Las proporciones de vacas cuyo estro duró menos de 8 h, de 8 a 16 h o más de 16 h fueron analizadas entre tratamientos, dentro de cada categoría, mediante una prueba de Chi cuadrada. Con el mismo método se analizó el efecto del tratamiento sobre la hora de inicio del estro, para lo cual se crearon cuatro categorías dentro de un periodo de 24 h (1:00-6:00, 6:01-12:00, 12:01-18:00 y 18:01-24:00) de acuerdo a lo establecido por White et al. (2002) y la comparación de las proporciones entre tratamientos se realizaron dentro de cada categoría. El número de montas ocurridas dentro de las categorías de horario anteriores fueron comparadas entre tratamientos mediante el siguiente modelo lineal general: donde
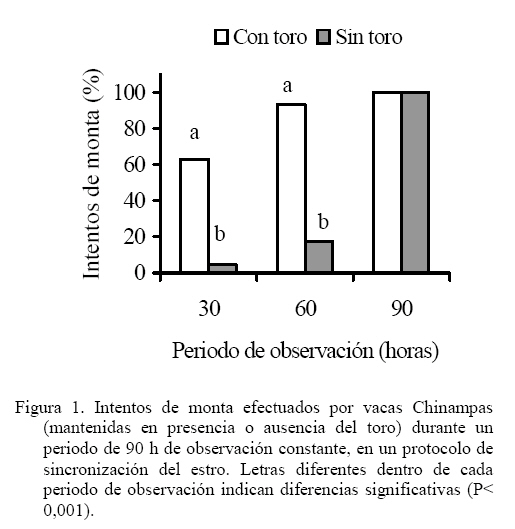
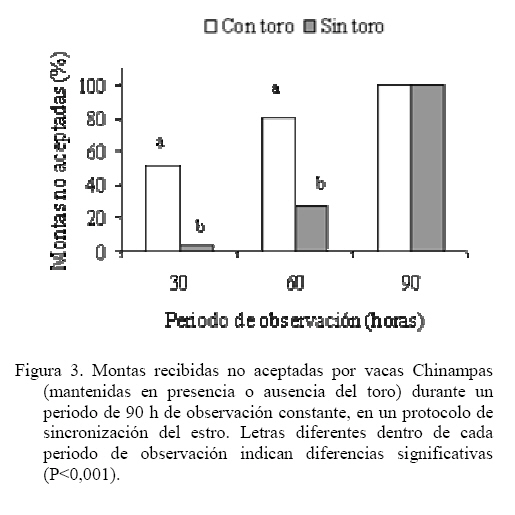
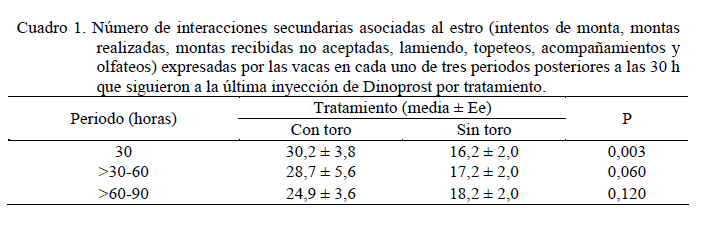
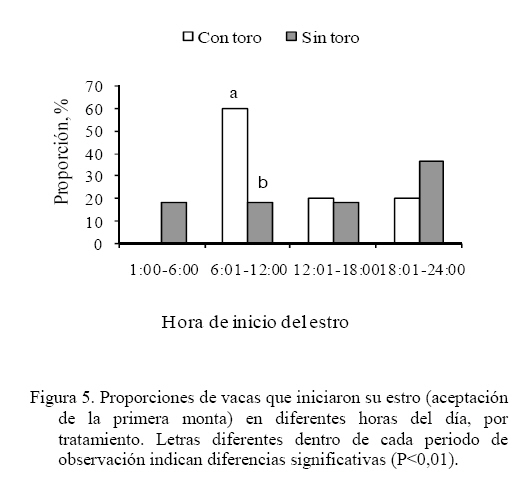
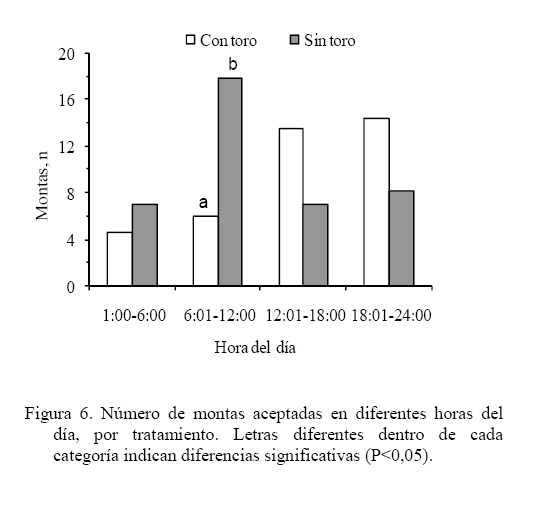
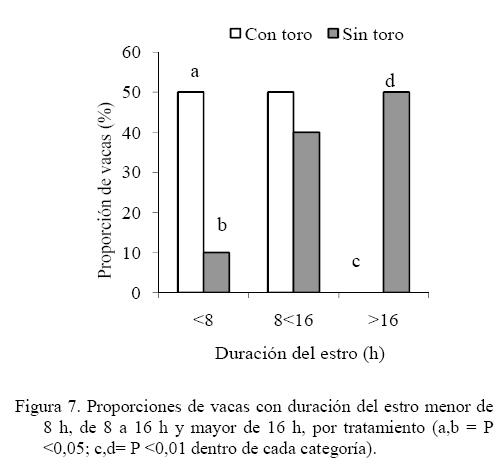
Las medias de los datos no categóricos se compararon mediante el método de diferencia mínima significativa (LSD). Todos los procedimientos estadísticos utilizados fueron desarrollados a través del paquete SAS (SAS, 1989). RESULTADOS Y DISCUSIÓN Comportamiento sexual Durante las 30 h que siguieron a la segunda inyección de Dinoprost no se registraron interacciones secundarias ni vacas en estro, por tal motivo la presentación y discusión de los resultados se hace sobre los acontecimientos que ocurrieron a partir de ese momento. En momentos variables, posteriores a esas 30 h, todas las vacas presentaron celo. Como se observa en la Figura 1, dentro de las primeras 30 y 60 h de observación constante se registró una mayor proporción de IM en las vacas que estuvieron con el toro. En las primeras 30 h se registró el 63% de los IM en G1 y el 5% en G2, diferencia que resulta altamente significativa (P<0,001). A las 60 h de observación constante se mantuvo la tendencia a un mayor registro de eventos (P<0,001) en G1 (93%) que en G2 (36,5%). La proporción de MR siguió la misma tendencia que los IM (Figura 2). En las primeras 30 h de observación constante se observaron más MR en G1 (43%) que en G2 (0%; P<0.001). Para las 60 h, se mantuvo la tendencia a un mayor registro de eventos (P<0,001) en G1 (84,5%) que en G2 (17,6%). En la Figura 3 se observa que la proporción de MRNA en las primeras 30 h de observación constante fue más alta en G1 (52%) que en G2 (4%; P<0,001). Para las 60 h, se mantuvo la misma tendencia a un mayor registro de eventos (P<0,001) en G1 (80,5%) que en G2 (27,3%). En la Figura 4 se aprecia que la proporción de MRA, las cuales indican un celo franco, fue de 51% en las primeras 30 h de observación constante en G1 y de 0% en G2, lo cual resultó en una diferencia altamente significativa (P<0,001). Para las 60 h, se mantuvo la tendencia a un mayor registro de eventos (P<0,001) en G1 (84,5%) que en G2 (14,5%). Un comportamiento similar de las interacciones sexuales fue reportado por Galina et al. (1987) en vacas cebú. Señalan que la presencia del toro cambia los patrones de conducta estral en el hato; en ese estudio, las vacas mantenidas en pradera mostraron actividad estral en diferentes momentos después de la inyección de Dinoprost dependiendo de la presencia o ausencia del macho. Al respecto, Chenoweth (2000) señala que los machos son capaces de inducir reflejos neuroendocrinos que alteran la función ovárica en hembras de la misma especie. El estímulo biológico que deriva del toro para alterar la actividad reproductiva de las vacas no se conoce. Algunos investigadores especulan que el toro produce una feromona iniciadora que actúa a través de una ruta olfatoria para provocar la respuesta (Rekwot et al., 2001). Barauh y Kanchev (1993) demostraron que la orina del toro asperjada dentro del conducto nasal de vacas lecheras 7 d después del parto incrementó la concentración sistémica de LH y FSH dentro de los primeros 70 minutos de la exposición. De la misma manera, Fernández et al. (1996) encontraron que las vacas en la etapa posparto expuestas de manera intermitente a toros incrementaron su concentración media de LH y la frecuencia de pulsos de la misma con cada exposición realizada a intervalos de 2 h. El 100% de las diferentes interacciones asociadas al comportamiento sexual en ambos grupos del presente estudio se registraron dentro de las 90 h de iniciada la observación permanente. La proporción de LAM, TOPE y OLF a las 30 h fue mayor (P<0.001) en G1 (25,9 28,9 y 33,8%, respectivamente) que en G2 (12,8 15,0 y 4,6%, respectivamente). Una tendencia similar (P<0,001) se mantuvo a las 60 h de observación continua (G1 = 78,5 70,0 y 73,0%; G2 = 60,0 32,0 y 23,0% para LAM, TOPE y OLF, respectivamente). En el Cuadro 1 se pueden observar el número de interacciones secundarias (IM, MR, MRNA, LAM, TOPE, ACOM y OLF) previas y posteriores a la aceptación de la monta en diferentes tiempos después de iniciada la observación permanente (30, > 30-60 y > 60-90 h). Dentro de las primeras 30 h se registraron más interacciones en G1 (P<0,01). Después de las 30 h, hasta las 60 h de observación hubo una tendencia (P<0,06) a una mayor cantidad de eventos en G1 y después de las 60 h hasta las 90 h la cantidad de interacciones registradas fue igual entre tratamientos (P>0,05). De manera similar, Silva et al. (2002) trabajando con vacas Brahman en un programa de sincronización del estro con un implante subcutáneo de progestágeno, consideran que la introducción del toro 40 h después del retiro de los implantes pudo haber contribuido al aumento del porcentaje de manifestación de estros que se observó entre las 40 y las 50 h. Resultados diferentes fueron constatados por Orihuela et al. (1983) en ganado cebú, quienes reportan que cuando las vacas estuvieron en ausencia del toro la mayoría de las interacciones sexuales se circunscribieron a un periodo entre 50 y 70 h después de la segunda inyección de PGF2α, y cuando el macho estuvo presente, la actividad se extendió hasta el final de un periodo de observación de 100 h. El total de MA por vaca fue similar entre grupos (39,0 ± 7,8 y 44,1 ± 9,5 en G1 y G2 respectivamente; P>0,05). Valores inferiores en invierno y verano (29 y 21, respectivamente) fueron obtenidos por Ávila et al. (2003) en vacas Gyr. En otros estudios se ha observado un menor número de montas aceptadas, como en el caso de las vaquillas Criollas Sanmartinero de Colombia con 14,5 montas (Góngora y Hernández, 2006) y de vacas Holstein x Brahman de Venezuela (2,2 montas) como lo describen Ramírez-Iglesia et al. (2002). Valores superiores (50 montas) han sido reportados en vaquillas para carne (Stevenson et al., 1996). También hay trabajos (White et al., 2002) en vacas para carne que reportan una cantidad de montas por estro ligeramente superior que las observadas en el presente estudio, constatándose también el efecto de la estación de año sobre dicha variable (59, 43 y 38 en invierno, verano y primavera, respectivamente). En vacas Holstein en pastoreo, el número de montas durante el estro ha variado entre 8 y 33 (Xu et al., 1998). La cantidad de montas aceptadas por vaca dentro de los periodos de 30, > 30-60 y > 60-90 h de observación constante se presentan en el Cuadro 2. Hasta las 30 h se registraron más MA en G1 (P<0,01). Después de las 30 h, hasta las 60 h, la cantidad de MA fue similar entre los tratamientos (P>0,05) y después de las 60 h de observación, hasta las 90 h se constataron más MA en las vacas que se mantuvieron aisladas del macho (P<0,01). Valores inferiores en vacas Cebú han sido constatados por Orihuela et al. (1983) quienes reportan 15,9 y 13,9 montas por hembra en estro, cuando permanecieron aisladas o en presencia del toro, respectivamente. En vaquillas Brahman en estro se han reportado de 13 a 41 montas homosexuales y de 2 a 10 montas realizadas por el macho cuando se hicieron las observaciones después de un programa de sincronización del estro con Crestar (Silva et al., 2002). Características del estro Las vacas que fueron cortejadas por lo menos por una compañera o el toro celador y mostraban interés por la monta, topeteos y seguimiento fueron consideradas en proestro, o en estro cuando permitían la monta. Se pudo observar que los signos considerados como indicadores del proestro se manifestaron escasamente hasta las 60 h en las vacas que se mantuvieron aisladas del macho. La explicación de lo anterior se podría basar en el hecho de que algunos investigadores especulan que el toro produce una feromona iniciadora que actúa a través de una ruta olfatoria para generar una respuesta (Rekwot et al., 2001). Distribución de las montas e intervalo entre montas Como se indica en la Figura 5, el 60% de las vacas del G1 iniciaron el estro entre las 6:01 y 12:00 h, lo que resultó superior (P<0,05) a la observada en G2 (18,2%) dentro del mismo periodo; el resto de los estros en G1 se iniciaron entre las 12:01 y las 24:00 h (40%). En el grupo G2, el 54,6% de los estros iniciaron durante la noche (entre las 18:00 y 6:00 h). Los resultados observados en el grupo G1 concuerdan con las observaciones reportadas por Diskin y Sreeman (2000) quienes reportan que el 40% de las vacas lecheras en pastoreo, aceptaron por primera vez la monta a las 7:00 h. A diferencia de lo anterior, Dransfield et al. (1998) encontraron que en las vacas Holstein, la hora de inicio del estro se distribuye de manera casi uniforme a lo largo de los periodos establecidos para el presente trabajo. El 60% de las montas aceptadas en las vacas de G1 se concentraron entre las 6:01 y 12:00 h, lo que resultó superior (P<0,01) a la observada en G2 (18,2%) dentro del mismo periodo. El resto de las montas aceptadas en las vacas de G1 se distribuyeron de manera uniforme entre las 12:01-18:00 y las 18:01-24:00 h y las proporciones fueron similares entre tratamientos (P>0,05). No obstante, la distribución del inicio de estros en G2 fue uniforme entre las cuatro categorías de horario. Estos resultados concuerdan con el estudio realizado por Góngora y Hernández (2006) quienes reportaron que el 72% de los estros observados en vaquillas Criollas Sanmartineras ocurrieron entre las 5:00 y 11:00 h. Observaciones similares fueron hechas por White et al. (2002) en vacas Hereford x Angus las cuales recibieron 3,2 montas por hora entre las 6:00 y 12:00 h y solo 2,1 montas por hora en las 18 h restantes. En otro estudio se observó que la mayor actividad de monta se registró entre 6:00 y 9:00 h en vacas que permanecieron aisladas (25%) o en presencia del toro (33%), respectivamente (Orihuela et al., 1983). En ese mismo trabajo, el 63% de todas las montas en las vacas aisladas del macho y el 53% de las que estuvieron con el toro se observaron durante la noche (18:00 a 6:00 h). De manera similar, en el presente estudio, el 72,8% de las vacas en G2, concentraron su actividad de monta en el horario antes señalado. En este trabajo, el intervalo promedio entre MA fue de 24,9 ± 5,6 y 24,6 ± 3,5 minutos para las vacas que permanecieron con el macho o aisladas del mismo, respectivamente (P>0,05). Intervalos mayores entre montas fueron observados por White et al. (2002) en vacas Hereford x Angus durante el verano (4,1 h) y la primavera o el invierno (2,7 h). Esas discrepancias de resultados con el presente estudio podrían derivar de algunos factores asociados con el manejo, ya que las vacas en este trabajo se mantuvieron en un corral para su observación, mientras que en el experimento de White et al. (2002), los animales estuvieron en una pradera de 12 ha. Por otro lado, en ese trabajo las vacas presentaron estro natural en cambio, en nuestro estudio los estros estuvieron sincronizados. Esos dos factores concentraron la actividad en el tiempo y en el espacio, lo cual pudo haber influido en la discrepancia de los resultados. Interacciones sexuales primarias y secundarias El número total de interacciones (195 ± 25 y 217 ± 30 en G1 y G2, respectivamente) y la cantidad de MA resultaron similares entre tratamientos (P>0,05) con tendencia de estas últimas a una mayor cantidad en las vacas que estuvieron aisladas del toro (39,1 ± 7,8 y 44,1 ± 9,5 para G1 y G2, respectivamente). Al respecto, Ramírez-Iglesia et al. (2000) señalan que, tanto en la interacción heterosexual como en la homosexual, se observaron los mismos signos (flehmen, lamidos y olfateos) y que dichas expresiones fueron más abundantes en la interacción vaca-toro. El número de interacciones previas a la aceptación de la monta fue diferente (P<0,01) entre G1 (5,6 ± 1,5) y G2 (31,8 ± 3,4). Sin embargo, la cantidad de dichos signos secundarios del estro fueron similares entre tratamientos (P>0,05) después de la última monta aceptada (4,0 ± 0,89 y 3,4 ± 1,0 para G1 y G2, respectivamente). En un estudio de Yoshida y Nakao (2005) con vacas Holstein, los animales mostraron signos secundarios de estro durante un periodo de 9,6 h antes del inicio y durante 18,4 h después de finalizado el mismo. En el presente trabajo, el número de montas aceptadas en el grupo de vacas que permanecieron con el macho tuvo una distribución similar (P>0,05) en las diferentes horas del día. Sin embargo, en las vacas que permanecieron aisladas del toro hubo más montas (P<0,05) entre las 6:01 y las 12:00 h (17,8 ± 4,5) que en el resto de las categorías (Figura 6). El menor número de interacciones registradas en las hembras mantenidas con el macho, podría deberse a que cuando un toro es introducido en un grupo de vacas sincronizadas, este provoca varios cambios en su comportamiento. El toro establece dominancia sobre las vacas más grandes (Galina et al., 1987) y la subordinación puede conducir a una atenuación del comportamiento sexual (Hafez y Hafez, 2000). Duración del estro La DE fue similar en las vacas que se mantuvieron con el toro (10,7 ± 1,1 h) a las del grupo testigo (16,3 ± 2,6 h), pero se observa una tendencia (P< 0,07) a una menor duración de la aceptación de la monta en las vacas de G1. Los resultados coinciden con un reporte de Vaca et al. (1985) quienes indican que la duración del estro en la hembra bovina se reduce con la presencia del toro. En otro estudio con vaquillas Criollas Sanmartinero de Colombia, se observo una duración del estro similar cuando estas fueron mantenidas con toros celadores en un programa de sincronización de estros con Norgéstomet y valerato de estradiol (Góngora y Hernández, 2006). En una investigación de Silva et al. (2002) con vaquillas Brahman en el trópico Mexicano utilizando sincronización del estro con Norgéstomet y valerato de estradiol, observaron una variación de 8 a 18 h en la duración del estro. Cabe destacar que en ese estudio las vaquillas fueron mantenidas con el macho a partir de las 40 h de haber retirado el implante de norgéstomet. Estros de 13,2 y 10,9 h durante el invierno y el verano, respectivamente han sido reportadas para vacas Gyr (Ávila et al., 2003). Duraciones de estro similares a las observadas en el grupo testigo del presente trabajo fueron establecidas por White et al. (2002) en vacas Hereford x Angus de Oklahoma durante el verano (17,6 h) y el invierno (15,5 h). En la Figura 7 se puede observar que la proporción de vacas con una duración del estro de menos de 8 h fue diferente entre tratamientos (50 y 10% para G1 y G2, respectivamente; P<0,05). El 50% de las vacas en G1 presentaron un estro con duración de 8 a 16 h, porcentaje similar al observado en G2 (40%) para la misma categoría (P>0,05). Lo anterior significa que el 100% de las vacas que estuvieron en presencia del macho tuvieron un celo con duración no mayor de 16 h. En la Figura 7 se aprecia también que el 50% de las vacas en G2 presentaron un celo que duró más de 16 h. El 90% de los estros en este grupo duraron más de 8 h. CONCLUSIONES Se concluye que en presencia del macho, las vacas Chinampas sincronizadas con Dinoprost (Lutalize) concentraron en un periodo más corto la manifestación de los eventos asociados a la conducta estral y aumentó la proporción de estros con una duración menor de 8 h. Además, la presencia del macho influye en la hora de inicio del estro, agrupó la mayor proporción de estros en ciertas horas del día pero no influyó en el número de montas aceptadas por vaca ni en la duración del estro. LITERATURA CITADA
Copyright 2007 - Zootecnia Tropical The following images related to this document are available:Photo images[zt07003f1.jpg] [zt07003t1.jpg] [zt07003f5.jpg] [zt07003t2.jpg] [zt07003f2.jpg] [zt07003f4.jpg] [zt07003f3.jpg] [zt07003f6.jpg] [zt07003f7.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}