 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
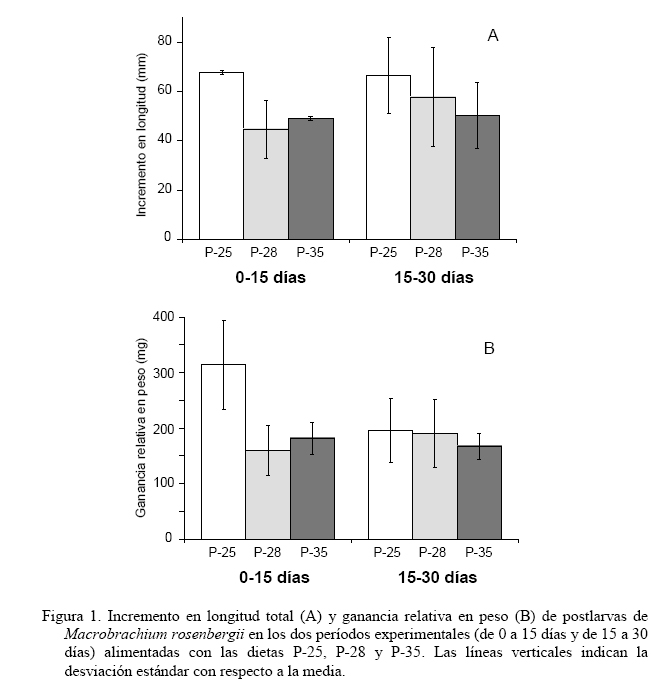
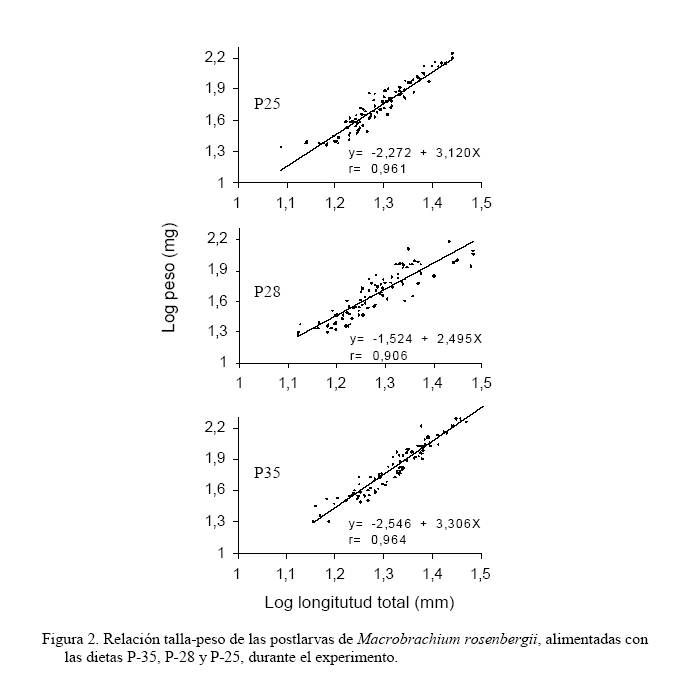
Zootecnia Tropical, Vol. 25, No. 2, 2007, pp. 111-121 Evaluación de tres dietas con diferente contenido proteico en el cultivo de postlarvas del langostino de río Macrobrachium rosenbergii Evaluation of three diets with different protein content in the culture of river prawn postlarvae, Macrobrachium rosenbergii Merirros Luna1, César Graziani1,4*, Elvis Villarroel1,4, Mairin Lemus2, César Lodeiros3,4 y Greys Salazar1 1 Lab. Camarones Dulceacuícolas, Dpto. Biología, Escuela de Ciencias, Núcleo Sucre, Universidad de Oriente, Cumaná 6101, Sucre. Venezuela. Code Number: zt07014 RESUMEN Se evaluó el crecimiento y la supervivencia de postlarvas de Macrobrachium rosenbergii alimentadas con dos dietas comerciales empleadas actualmente en cultivo de peces dulceacuícolas (P-28 y P-25) y una dieta formulada para camarones marinos (P-35) con distintos niveles de proteínas. El ensayo tuvo una duración de 30 días, durante los cuales se realizaron dos muestreos quincenales. Las postlarvas fueron distribuidas a una densidad de 50 organismos por acuario de 3,2 L, empleándose un total de cuatro réplicas experimentales y una de sustitución para cada dieta, con un suministro de alimento dos veces al día, equivalente al 10% de la biomasa húmeda de los camarones. La relación talla-peso de los organismos de las diferentes dietas mostraron relaciones lineales y significativas con un elevado índice de correlación (r>0,90) y un crecimiento alométrico, sin obtener diferencias significativas entre sus pendientes, mostrando que las dietas utilizadas fueron adecuadas para el desarrollo de los organismos. No obstante, en los demás parámetros de crecimiento se encontraron diferencias significativas entre las dietas utilizadas, particularmente a los 15 días del experimento, donde la dieta P-25 mostró el valor mayor en cuanto a longitud total y ganancia relativa en peso. El índice de ARN/ADN al día 30 mostró ser más elevado para la dieta P-25 y P-35 y ésta igual a la P-28. Al final del estudio, la tasa de conversión alimentaria de la dieta P-25 (1,57) fue menor, en un orden de más del 27% que las demás dietas. Estos resultados, y los análisis bioquímicos de las dietas evaluadas sugieren la utilización de dietas con un 25% de proteínas y elevado contenido de carbohidratos, para la alimentación de postlarvas de M. rosenbergii, recomendándose ensayar dietas con niveles proteicos inferiores a 25%, con la finalidad de verificar si son convenientes en función de disminuir aún más los costos del alimento. Palabras clave: Marobrachium rosenbergii, dietas, contenido proteico, postlarvas, langostino. ABSTRACT We evaluated the growth and survival of M. rosenbergii postlarvae fed with two commercial diets currently used for culturing freshwater fishes (P-28 and P-25) and a diet formulated for marine shrimp culture (P35), having different protein levels. The experiment lasted 30 days carrying out sampling of the animals after 15 and 30 days of culture. The postlarvae were cultured in 3.2 L aquaria at a density of 50 organisms per aquaria, using four replicates and one aquarium for replacements, for each diet. The diets were administered twice a day at a rate of about 10% of wet body mass of the animals. The relationship length/weight of the animals with the different diets were significant with a high correlation coefficient (r>0.90) and followed a linear model. They showed an alometric growth, and non significant differences among their slopes. Other growth parameters did show statistically significant differences among the diets, especially at 15 days of culture. At this time, P-25 showed highest total length and relative weight gain. The RNA/DNA index at day 30 was highest for P-25, and P-35 was intermediate. At this time, food conversion ratio for P-25 (1.57) was about 27% lower than the other two diets. These results, along with the biochemical analysis of the evaluated diets, suggest the use of diets with 25% protein content and high carbohydrate content for culture of M. rosenbergii postlarvae. It is proposed to study the use of diets with protein content lower than 25% to determine the possibility to lower the costs. Keywords: Macrobrachium rosenbergii, diet, protein content, postlarvae, prawn. INTRODUCCIÓN Los camarones dulceacuícolas del género Macrobrachium, familia Palaemonidae, son los comercialmente más importantes en esta familia. Las más de 100 especies pertenecientes a la familia se encuentran distribuidas en las zonas tropicales y subtropicales del mundo, siendo Macrobrachium rosenbergii la que ha sido cultivada con mayor éxito en diversos países del mundo, incluyendo las regiones tropicales del Caribe (New y Singholka, 1984; Valenti, 2006). La producción de postlatvas de M. roserbergii de buena calidad es importante para el éxito del cultivo, por lo que estudios de dietas que cubran eficientemente sus requerimientos energéticos son de carácter fundamental. Existen algunos estudios nutricionales en estas fases, en los cuales se han evaluado aspectos como niveles y fuentes de las proteínas, lípidos y carbohidratos utilizados en las dietas y su efecto en el crecimiento, supervivencia, tasa de conversión, asimilación, producción y costos operacionales (Molina-Vozzo et al., 1995; Lobão et al., 1995a,b; Correia et al., 1997), resaltando a las proteínas como el principal componente de las dietas comerciales, debido a la gran influencia en el crecimiento y su relación directa con los costos de producción (Cortés-Jacinto et al., 2003, Teshima et al., 2006). En Venezuela, actualmente se ofertan en el mercado algunos alimentos para especies en cultivo, de variadas formas y composiciones, que no se conoce si satisfacen los requerimientos nutricionales de los camarones de agua dulce, ya que han sido formuladas principalmente para camarones marinos y peces de agua dulce. Debido a ello, el presente trabajo evalúa el efecto de tres dietas comerciales con diferentes concentraciones proteicas en el desarrollo de postlarvas de Macrobrachium rosenbergii. MATERIALES Y MÉTODOS Las postlarvas (PL) de Macrobrachium rosenbergii fueron producidas en el Laboratorio de Camarones Dulceacuícolas de la Universidad de Oriente, siguiendo los protocolos estándares desarrollados por New y Singholka (1984). Para la experimentación, las postlarvas producidas se mantuvieron en acuarios, con un volumen de 3,2 L y un área de fondo de 0,13 m2, a una densidad de cultivo de 600 PL/m2, menor a la establecida por New y Singholka (1984) en protocolos estándares para su acopio (1.000 PL/m2), en función de minimizar el efecto de la densidad como factor perturbador. De acuerdo a ello, se colocaron 50 PL/acuario, empleándose cuatro réplicas experimentales por dieta y una de reposición. Tanto las PL de las réplicas de experimentación como las de sustitución, en todas las dietas, fueron del mismo tamaño y peso (ANOVA, P>0,1) con promedios generales de 11,6±1,37 mg y 12,02±0,45 mm, respectivamente. Durante el bioensayo la temperatura se mantuvo constante a 28±1°C; para ello todos los acuarios réplicas estuvieron semisumergidos en un recipiente con agua provisto de calentadores automáticos y un agitador para la homegeneización del agua. Todos los acuarios estuvieron provistos de aireación constante y se realizaron recambios diarios del 50% del agua para garantizar la calidad de la misma. Se ensayaron tres piensos comerciales, los cuales por acuerdo con la empresa transnacional que los suministró, se decidió identificar como P-35, P-28 y P-25 (el número corresponde al nivel proteico, en porcentaje, de la dieta) utilizados para la alimentación de camarones marinos y peces de agua dulce conocidos como tilapias (Oreochromis spp.), cachamas (Colossoma macropomum) y morocotos (Piaractus brachipomus), respectivamente. A cada alimento ensayado, se le determinó el porcentaje de cenizas (Covenin, 1979), carbohidratos (Chávez y González, 1995) y el contenido calórico (García, 1987). El alimento se suministró, en un equivalente al 10% de la biomasa húmeda en dos raciones diarias (mañana y tarde), durante 30 días. El tamaño del "pellet" de los tres alimentos se ajustó, mediante macerado y tamizado a 1 mm de longitud. En todos los acuarios, diariamente se contabilizó el número de postlarvas vivas en función de determinar la sobrevivencia. Quincenalmente se tomaron cuatro PL por réplica de tratamiento, las cuales fueron inmediatamente sustituidas por postlarvas de los acuarios de reposición, las cuales mantenían las mismas condiciones, con la finalidad de no afectar la densidad. Las cuatro postlarvas fueron utilizadas para formar un "pool" por réplica. A cada "pool" se le determinó el peso en húmedo y se llevó a una estufa por 48 horas a una temperatura de 60ºC, para obtener el peso en seco. Posteriormente, las muestras fueron colocadas en una mufla a 500ºC durante tres horas. Por diferencia entre el peso en seco y el de las cenizas, se obtuvo la materia orgánica libre de cenizas, masa que fue utilizada para realizar los diversos índices gravimétricos. Para evaluar las dietas se establecieron varios índices, determinando la longitud total de las postlarvas con un vernier (0,01 mm de precisión) y sus pesos con una balanza analítica (0,0001 g de precisión), de esta manera se estableció la ganancia relativa en peso (GRP) definida como la relación en porcentaje del incremento del peso con respecto al peso al inicio del experimento, la tasa de conversión alimentaria (TCA) en relación del peso en seco del alimento sobre el incremento del peso en seco de las postlarvas. De igual manera, se determinó el crecimiento instantáneo mediante la relación de los ácidos nucleicos ARN/ADN, determinados por la técnica de fluorescencia de Canino y Calderone (1995). Para ello, se utilizaron en cada muestreo otras cuatro postlarvas, de igual manera que para obtener los diferentes pesos descritos anteriormente. Para determinar el efecto de las dietas en las variables analizadas en las postlarvas de M. rosenbergii, en cada período experimental y previa comprobación de la normalidad de los datos y la homegeneidad de las varianzas, se realizó un análisis de varianza simple. Las diferencias entre las dietas se establecieron con la prueba a posteriori de Duncan (Steel y Torrie, 1985). Además, se realizó un análisis de regresión para establecer el grado de relación entre el peso y la talla, previa transformación logarítmica de los datos para cada dieta y se realizaron análisis de comparación de las pendientes mediante el contraste en pares, siguiendo las recomendaciones de Zar (1984). RESULTADOS El Cuadro 1 muestra la composición nutricional y energética de las distintas dietas empleadas. Además de la proporción del contenido de proteínas, las diferencias notables de las macromoléculas entre las dietas fue fundamentalmente de carbohidratos, donde la dieta P-25 posee una elevada proporción de casi 50%, en comparación con 38% para P-35 y 31% para P-28. La cantidad de grasas, en todas las dietas, fue la de menor proporción entre las macromoléculas (<5%), siendo la dieta P-25 la que mostró menor concentración (2,2%). De esta manera, la energía en kcal/g de las dietas fue mayor en la P-35 (357), seguido de la P-25 (335) y la P-28 (288). Una diferencia notable también se estableció en el contenido de fibras, donde la dieta P-28 superó en más del doble a las demás dietas. El incremento en longitud total mostró diferencias significativas entre las dietas en el período inicial (0-15 días, P<0,05); sin embargo, no para el período final (15 a 30 días, P>0,05). En ambos períodos, los organismos alimentados con la dieta P-25 fueron los que más incrementaron en longitud, siendo estos incrementos significativos (P<0,05) sólo en el período inicial, donde las dietas P-28 y P-35 no mostraron diferencias significativas (P>0,05) (Figura 1 A). Esta misma tendencia, en cuanto a la diferenciación de los efectos de las dietas se desarrolló para la ganancia relativa en peso (Figura 1 B). Las relaciones talla-peso de las diferentes dietas (Figura 2) resultaron ser lineales significativamente (P<0.001), con coeficientes de correlación elevados (r>0,90) y pendientes que muestran un crecimiento alométrico, siendo la de la dieta P-25 la más elevada, aunque no significativamente diferente de las demás (comparación múltiple entre pendientes (P<0,05).
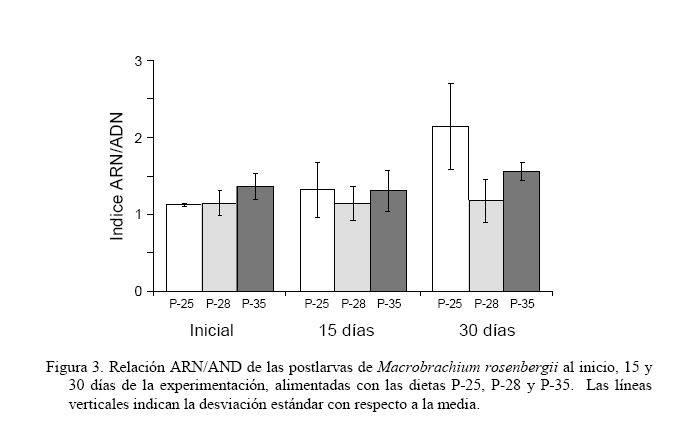
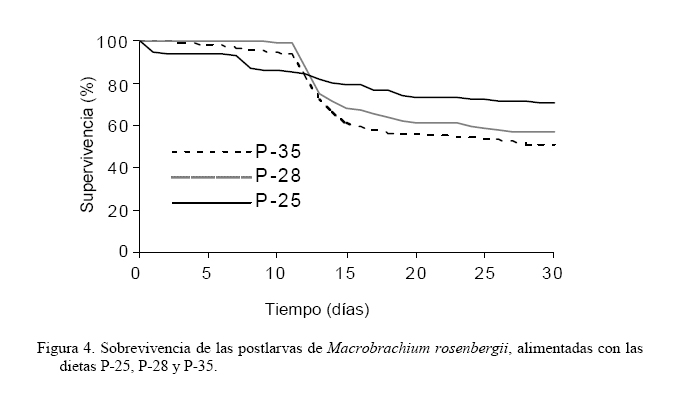
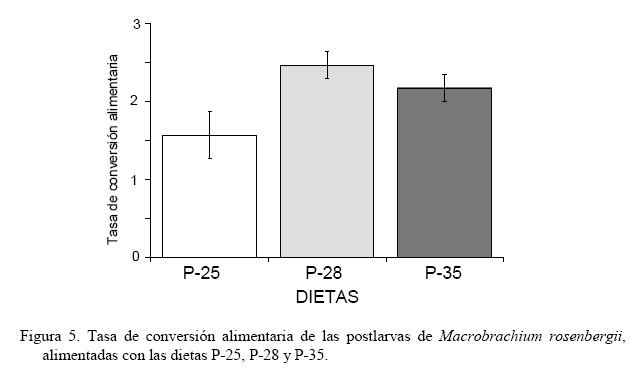
El índice de crecimiento instantáneo, relación ARN/ADN (Figura 3), mostró valores significativamente iguales (P>0,05) al inicio y a los 15 días del experimento; no obstante, al final del estudio la dieta P-25 fue la que obtuvo el mayor valor 2,14±0,055 (P<0,05), aunque significativamente igual a la dieta P-35 (1,56±0,119) y ésta a la dieta P-28 con un índice de 1,18±0,283 (P>0,05). Durante los primeros 12 días del ensayo, la mayor supervivencia se obtuvo con las dietas P-28 y P-35 (>95%), encontrándose para P-25 valores entre 85-95% (Figura 4). A partir de ese momento, y en 5 días, para estas dietas, la supervivencia decayó significativamente a 60-70% (P<0,05) para luego estabilizarse, sin cambios buscos, obteniéndose al final del experimento supervivencias entre el 50-65%. En contraste, la dieta P-25, aunque durante la primera semana presentó disminución de la supervivencia, mostró menores tasas de mortalidad a través del período experimental, alcanzando una supervivencia, al final del experimento, por encima del 70%. Estos valores permitieron establecer que la dieta P-25 fue la que obtuvo significativamente (P<0,05) mayor supervivencia. Al final del experimento la dieta P-25 generó una tasa de conversión alimenticia (TCA, Figura 5) de 1,57±0,299, significativamente menor (P<0,05) que las obtenidas con las dietas P-35 (2,17±0,174) y P-28 (2,46±0,170). DISCUSIÓN Con todas las dietas se observó un crecimiento alométrico, sin diferencias significativas en la relación talla-peso, sugiriendo que todas las dietas utilizadas fueron adecuadas para el desarrollo de los organismos. Sin embargo, los organismos alimentados con las dietas mostraron diferencias en los parámetros de crecimiento evaluados y la supervivencia, indicando diferencias en cuanto a la asimilación del alimento. Estas diferencias mostraron a la dieta P-25 con los valores más adecuados para el desarrollo y crecimiento de las postlarvas de M. rosenbergii, lo cual fue corroborado con la TCA, cuyo valor medio alcanzado (1,57) fue menor en más del 27% de las demás dietas. Estas diferencias podrían estar asociadas al contenido proteico de la dieta (25%) sosteniendo resultados obtenidos en otros trabajos (Sandifer y Joseph, 1976; Stanley y Moore, 1983), los cuales han señalado niveles de 25% de proteínas como óptimo para el crecimiento de M. rosenbergii. No obstante, algunos estudios no muestran diferencias en el crecimiento y supervivencia de juveniles de M. rosenberguii en rangos de 15-35% de proteínas (Balazs y Ross, 1976; Boonyaratpalin y New, 1980) y otros han señalado buenos resultados con niveles entre 40-50% (Molina-Vozzo et al., 1995) y 35% (El-Sayed, 1997). Algo similar, en cuanto a la variabilidad de resultados, ocurre con dietas probadas en camarones marinos, llegándose inclusive a recomendar para postlarvas dietas con proporciones de proteínas menores, como por ejemplo, Velasco et al. (2000) quienes proponen dietas entre 18 y 20% para Litopenaeus vannamei. Estas diferencias en resultados son probablemente producto del origen de los componentes proteicos y de los otros constituyentes de las dietas, así como del procesado de las dietas y de las condiciones de cultivo, lo cual pone de manifiesto la necesidad de profundizar en aspectos nutricionales y homogeneizar en lo posible los diseños experimentales, en función de establecer comparaciones entre los resultados con un grado de confiabilidad adecuado. Teniendo en cuenta los diferentes resultados anteriores y el del presente trabajo, aún cuando no se ensayaron dietas con niveles proteicos menores a 25%, no se justificaría dar un nivel más elevado de proteína si se puede abaratar los costos y obtener buenos resultados con niveles proteicos menores. De hecho, una proyección de costos de las dietas utilizadas en este estudio muestran que considerando la TCA final y los costos del alimento, la dieta P-25 representó un ahorro de aproximadamente 55% sobre el empleo del alimento P-35 formulado para camarones marinos y que actualmente se emplea en camarones dulceacuícolas. En el caso de la dieta P-28, a pesar de que fue 38% más costosa que la dieta P-25, su elevada TCA no la hizo igual de atractiva. Al considerar el período de experimentación en dos intervalos de cultivo, se encontró que durante el período final (15-30 días) se observaron los mayores incrementos en biomasa. Esto pudiera deberse a que durante el período inicial (0-15 días) del experimento, se observó una mayor frecuencia en las mudas, lo cual pudo conducir a canalizar la producción energética para el proceso de muda, restando ganancia en peso por crecimiento. A este respecto, Graziani et al. (1998) indicaron que durante las primeras fases de vida, las mudas en Macrobrachium spp. son frecuentes y por ello tienen que destinar gran parte de la energía a la formación del nuevo exoesqueleto. De esta manera, algunos estudios (Vega-Villasante et al., 2000; Molina et al., 2000) evidencian una ausencia en el consumo de alimentos, durante el cual, el camarón utiliza las reservas acumuladas en el hepatopáncreas para la construcción del nuevo exoesqueleto y en la síntesis de nuevo tejido relacionado con la muda. Una hipótesis adicional, a la poca ganancia en peso relacionado con la muda, es que en dichos procesos fisiológicos la alimentación podría verse afectada debido a que las estructuras quitinosas como mandíbulas, maxilas y dientes del molinillo gástrico de los camarones por estar recién generadas y blandas dejan de ser funcionales, ocasionando una limitante en la articulación mecánica para el proceso de alimentación. De esta manera, en la medida que las postlarvas son mayores y las mudas durante su desarrollo se distancian en el tiempo, la energía es más canalizada hacia el crecimiento. En nuestro estudio, la implicación de la muda como un proceso crítico en el crecimiento de las postlarvas pudo haberse reflejado en el período inicial, teniendo repercusión en la supervivencia. En las postlarvas alimentadas tanto con P-28 y P-35 se observó un aumento brusco de la mortalidad entre los días 12 y 14 de la experiencia, mientras que la mortalidad en las postlarvas alimentadas con P-25 fue baja sin diferencias notables. Aunque en este trabajo no se contabilizó el número de mudas, fue evidente el aumento en el número de mudas ocurridas entre los días 11 y 14 del cultivo en las postlarvas alimentadas con P-28 y P-35 (observación personal), lo que pudo ocasionar el aumento de la mortalidad. Además de las implicaciones antes señalas para el crecimiento, los camarones al mudar quedan con el exoesqueleto blando, haciéndose más susceptibles al canibalismo propio del género Macrobrachium (Graziani et al., 1993, 1998). A pesar que la concentración de proteínas en las dietas para organismos acuáticos son la principal atención de los investigadores debido a su influencia en el costo (New, 1976) y la síntesis y reparación de tejidos (Lim y Akiyama, 1992), la eficiencia de una dieta no depende únicamente de las proteínas. Bautista y Subosa (1997) señalan que los niveles de proteínas mejoran el crecimiento cuando se combinan con carbohidratos, lo cual supone una forma más económica de cubrir los requerimientos energéticos. En tal sentido, los carbohidratos, en las dietas para crustáceos derivados básicamente de harinas vegetales (maíz, trigo, arroz y sorgo) son fundamentales, ya que sus componentes son utilizados como fuente de energía, en la síntesis de quitina (principal constituyente del exoesqueleto) y en la formación de componentes fundamentales como ácidos grasos y esteroles (Clifford y Brick, 1978; Kucharski y Da Silva, 1991; Cruz-Suárez, 1996). De hecho, los carbohidratos son reportados por Díaz-Herrera et al. (1992) como el principal sustrato energético para cubrir los requerimientos metabólicos en postlarvas y juveniles de M. rosenbergii. Nuestros resultados soportan los estudios anteriores, ya que la dieta P-25% que proporcionó mayor crecimiento y supervivencia, así como menor TCA, fue la dieta con mayor proporción de carbohidratos con casi un 50% de su constitución macromolecular, a diferencia de las otra dietas (31% para P-35 y 38% para P-28). De esta manera, es muy probable que el papel y la eficiente utilización de la proteína estén siendo influenciados por el complemento de carbohidratos. En este sentido, Rosas et al. (2000) señalaron la posibilidad de reducir las proteínas de la dieta sustituyéndolas por carbohidratos, los cuales son mejor asimilados si se incluyen en forma de almidón. En esta investigación se empleó la relación ARN/ADN como índice de condición fisiológica. Este índice está siendo utilizado para indicar crecimiento instantáneo, o condición fisiológica en relación con la influencia ambiental o deficiencias en la alimentación en invertebrados (Lodeiros et al., 1996; Núñez et al., 2002). En nuestro estudio, el índice mostró valores iguales al inicio del experimento, corroborando la equidad en la condición inicial de los organismos, y tan solo mostró valores significativamente mayores al final del experimento, mostrando el valor mayor para la dieta P-25 (2,14), el cual supuso más del 80% del índice de las otras dietas, aunque las diferentes dietas mostraron índices elevados de dispersión, que la hizo significativamente igual al de la dieta P-35 (1,56) y ésta a la de la P-28 (1,18). La relación ARN/ADN, a pesar de ser tomada como un índice de condición que demuestra actividad celular de forma catabólica e instantánea debido a la actividad enzimática relativa al ARN en base al contenido celular estático de ADN y relacionado con el crecimiento instantáneo, debe ser tomado con mucha precaución, debido a que este parámetro no solo puede ser variable para el proceso de crecimiento, sino para otros procesos metabólicos celulares, sean de índole catabólico o anabólico, pudiendo enmascarar su relación con el crecimiento, como los procesos de demanda metabólica de la reproducción o elevada actividad por defensa ante estresores contaminantes (Lodeiros et al., 1996; Acosta y Lodeiros, 2001). En lo particular, y aunque los resultados del índice mostraron cierta correlatividad con los obtenidos con otros parámetros de crecimientos evaluados en el presente estudio, debido a los procesos de muda y las implicaciones metabólicas que éstos poseen en crustáceos, pensamos que el índice no debe ser aconsejable para evaluar condición fisiológica para estos organismos. Los resultados de este estudio, y las discrepancias encontradas con otros trabajos, ponen de manifiesto la variabilidad de los resultados y la necesidad de seguir evaluando alimentos comerciales disponibles en el mercado. Si bien falta mucho por investigar en cuanto a la nutrición de Macrobrachium, nuestro estudio propone la utilización de dietas con un 25% de proteínas y elevado contenido de carbohidratos. En este sentido, la utilización de los ensilados de maíz, son una materia prima utilizable, la cual inclusive puede sustituir el maíz, cumpliendo de esta manera con elevado contenido de carbohidratos y a la vez disminuyendo costos (Coelho-Emerenciano y Massamitu-Furuya, 2006). Independientemente de ello, es conveniente ensayar dietas con niveles proteicos inferiores a los empleados en este estudio, con la finalidad de verificar si son convenientes en M. rosenbergii, y para disminuir aún más los costos del alimento. En vista que la dieta P-25 arrojó los mejores resultados y es empleada actualmente en el cultivo de peces continentales, la hace ideal para ser utilizada en el policultivo de M. rosenbergii con otros peces, como una manera de aumentar la productividad y simplificar el manejo en el cultivo, al utilizar el mismo alimento para peces y camarones. No obstante, es recomendable realizar estudios de policultivo analizando factores como el tamaño de las postlarvas y/o alevines a sembrar y densidades, como una manera de minimizar impactos negativos en el crecimiento y sobrevivencia de ambas especies al competir por espacio y alimento. AGRADECIMIENTOS La presente investigación ha sido financiada parcialmente por el Consejo de Investigación de la Universidad de Oriente, Fundacite Anzoátegui y la Fundación para la Investigación y Desarrollo de la Acuicultura del estado Sucre (Fidaes) de la Gobernación del estado Sucre, Venezuela, siendo ésta su Contribución No. 1. LITERATURA CITADA
Copyright 2007 - Zootecnia Tropical The following images related to this document are available:Photo images[zt07014f5.jpg] [zt07014f4.jpg] [zt07014f3.jpg] [zt07014f2.jpg] [zt07014f1.jpg] | ||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}