 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
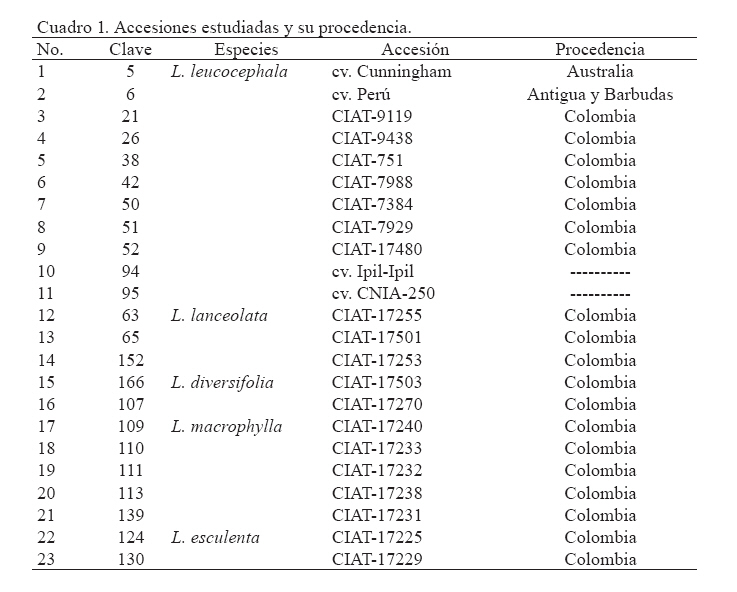
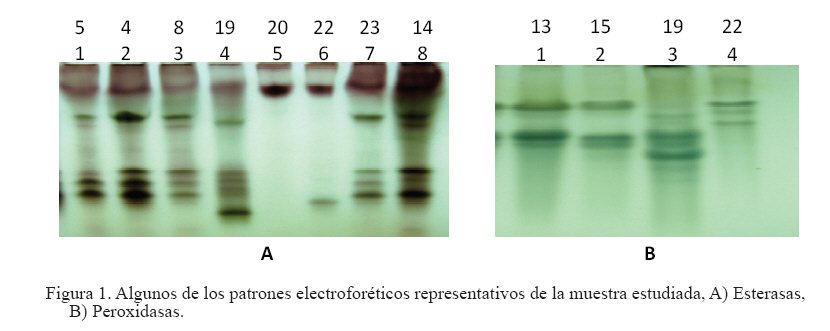
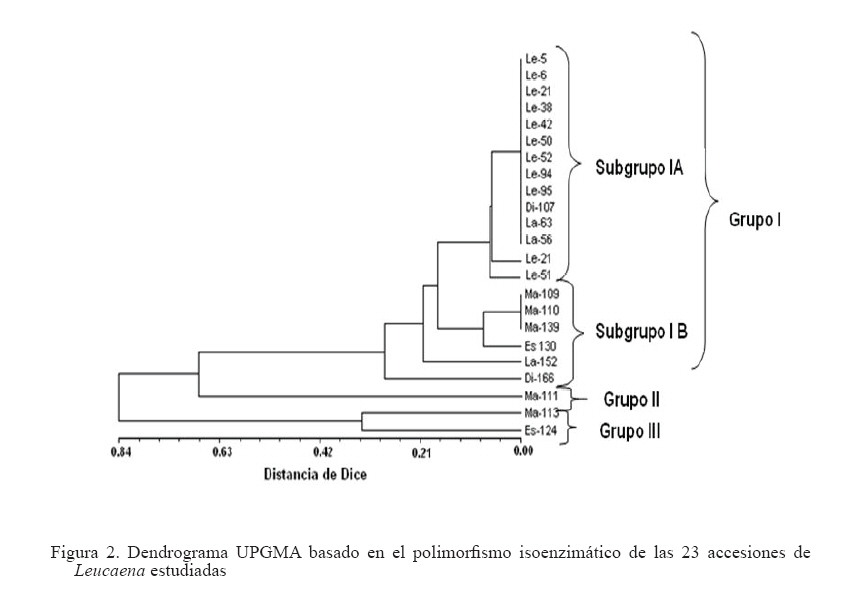
Zootecnia Tropical, Vol. 26, No. 3, 2008, pp. 327-331 Estudio del polimorfismo isoenzimático en el género Leucaena Study of isoenzymatic polymorphism in the Leucaena genus Hilda B. Wencomo1*, Maykelis Díaz1, Alba Álvarez2, Leonor Mora2 y Orlando Coto3 1 Estación Experimental de Pastos y Forrajes “Indio Hatuey”. Central España Republicana, Perico, Matanzas, Cuba. *Correo Code Number: zt08057 RESUMEN El trabajo se desarrolló con el objetivo de estudiar el polimorfismo isoenzimático de 23 accesiones del género Leucaena. También se incluyeron cinco especies de leucaena (L. leucocephala, L. lanceolata, L. diversifolia, L. macrophylla y L. esculenta), además de los cultivares comerciales L. leucocephala cv. Cunningham, L. leucocephala cv. Perú, L. leucocephala cv.Ipil-Ipil y L. leucocephala cv. CNIA-250. Para su desarrollo, se utilizaron los sistemas isoenzimáticos peroxidasas, α- y β- esterasas, malato deshidrogenasa y alcohol deshidrogenasas. Los datos obtenidos fueron procesados a través del análisis estadístico multivariado. Se pudo detectar la existencia de variabilidad genética dentro de la colección. Los sistemas isoenzimáticos (esterasas y peroxidasas) resultaron polimórficos en la muestra estudiada, principalmente las esterasas. Por otra parte, el análisis de diversidad genética permitió diferenciar L. leucocephala con una mayor claridad con respecto al resto de las especies estudiadas, aunque no se ganó en discriminación dentro de la especie. Los sistemas isoenzimáticos permitieron detectar diferencias entre las accesiones, siendo las esterasas las más polimórficas. Palabras clave: Leucaena, recursos genéticos, variabilidad, isoenzimas ABSTRACT This work was carried out with the objective of performing the izoenzymatic characterization of 23 accessions of Leucaena spp., which included five species of the Leucaena genus (L. leucocephala, L. lanceolata, L. diversifolia, L. macrophylla and L. esculenta), in addition to the commercial cultivars L. leucocephala cv. Cunningham, L. leucocephala cv. Perú, L. leucocephala cv. Ipil-Ipil, and L. leucocephala cv. CNIA-250. For the development of the trial, the isoenzyme analyses of peroxidases, α-and β-esterases, malato dehydrogenases, and alcohol dehydrogenases were made. The data obtained were subject to multivariate statistical analysis. The isoenzymatic systems (esterases and peroxidases) turned out to be polymorphic in the sample studied, mainly esterases, and the analysis of genetic diversity allowed to differentiate L. leucocephala with higher clarity than the rest of the species studied, although there was no gain regarding discrimination within the species. Keywords: Leucaena, genetic resources, variability, isoenzymes INTRODUCCIÓN En Cuba, los principales estudios de caracterización e identificación del género Leucaena especialmente de la especie L. leucocephala, se han centrado fundamentalmente en caracteres morfoagronómicos bajo condiciones de campo o en experimentos aislados, incluyendo solamente las cuatro variedades registradas como comerciales (Ipil-Ipil, Perú, Cunningham y CNIA-250) pertenecientes a la especie L. leucocephala. Sin embargo, hasta el momento no se han realizado estudios de caracterización más detallados que incluyan los marcadores isoenzimáticos, así como un mayor número de especies de este género. La caracterización de la diversidad genética, mediante técnicas bioquímicas como la electroforesis (de proteínas o isoenzimas) juega un papel importante como complemento de la caracterización morfoagronómica, al igual que el análisis directo de ADN (Schmidt et al., 2003). La electroforesis de isoenzimas favorece el empleo de marcadores genéticos más eficientes que los morfológicos en muchas ocasiones, a pesar de estar influidos por la acción ambiental y depender del tejido y estadio de desarrollo de la planta que se evalúa. Esta técnica es relativamente sencilla, poco costosa y codominante y permite distinguir los genotipos homocigóticos y heterocigóticos y desarrollar por tanto estudios de mapeo y ligamiento, de genética poblacional, entre otros. Atendiendo a lo anteriormente mencionado en el presente trabajo se desarrolló la caracterización isoenzimática de una muestra de 23 accesiones que incluyó cinco especies del género Leucaena (L. leucocephala, L. lanceolata, L. diversifolia, L. macrophylla y L. esculenta). MATERIALES Y MÉTODOS Se utilizaron rebrotes nuevos de las hojas de cada una de las accesiones evaluadas (Cuadro 1). Se maceraron 0,5 g del tejido foliar en nitrógeno líquido y se añadió buffer de extracción 1/1 (m/V). El extracto fue centrifugado a 14.000 rpm durante tres minutos y se colectó el sobrenadante para la electroforesis. Se empleó un buffer tris-citrato pH 8,3 al que se añadió KCl 0,08%, MgCl2 0,2%, EDTA 0,04% 0,5 mL de Tritón X100 1%, 2 mL 10% DTT y 25 mg PVP-40 4%. Las corridas se efectuaron en cámaras de electroforesis vertical (Modelo V16) y geles de poliacrilamida (8,5% para peroxidasas y alcohol deshidrogenasas y 12% para esterasas), a 4oC, 120 V, 20 mA, durante cuatro horas. En todas las corridas se añadieron 30 μL de cada una de las muestras y 10 μL de buffer de carga (BC). Se realizaron las tinciones específicas para peroxidasas (Prx, EC. 1.11.1.7), α- y β-esterasas (Est, EC. 3.1.1), malato deshidrogenasa (Mdh, EC. 1.1.1.37) y alcohol deshidrogenasas (Adh, EC. 1.1.1.1), según Álvarez et al. (2000). Las corridas electroforéticas se repitieron al menos tres veces y solo se registraron las bandas consistentes y reproducibles. Los fenotipos isoenzimáticos de cada accesión se registraron como presencia/ausencia de cada banda (0/1, respectivamente). La matriz binaria de datos isoenzimáticos (eliminando las bandas redundantes) se utilizó para generar una matriz de distancias genéticas entre todos los pares de genotipos, expresada como el complemento del coeficiente de Dice (Dice, 1945), usando el programa SIMQUAL del paquete estadístico NTSYS-pc versión 2 (NTSYS-pc, 1997). Se desarrolló un análisis de conglomerados, basado en la matriz de distancia de Dice. Para ello, se generó un dendrograma en el programa SHAN, del mismo paquete estadístico. El criterio de agregación utilizado fue el método de la media aritmética de grupos no ponderada UPGMA. Además se determinó la correlación cofenética (r), para determinar la homogeneidad entre la matriz de distancia y el dendrograma y para representar la exactitud de la técnica utilizada (NTSYS-pc, 1997). RESULTADOS Y DISCUSIÓN De los cinco sistemas isoenzimáticos analizados, peroxidadas, α- y β-esterasas, malato deshidrogenasa y alcohol deshidrogenasas, los tres primeros fueron polimórficos, a diferencia de los dos últimos que fueron monomórficos. En la Figura 1 se muestra los perfiles electroforéticos representativos de la muestra analizada para los sistemas Est y Prx. Como se puede apreciar a partir de estos resultados, las Est resultaron más polimórficas en cuanto a número de bandas polimórficas que permitieron detectar, el número de patrones electroforéticos y la frecuencia de los mismos en la muestra, en correspondencia con lo reportado acerca del alto polimorfismo de este sistema para numerosas especies vegetales (Schmidt et al., 2003). Las Prx dos zonas principales de actividad enzimática (Figura también tuvieron un alto nivel de polimorfismo en 1B), en correspondencia con lo reportado por Harris et cuanto a porcentaje de bandas polimórficas, aunque al. (1994) para L. leucocephala y a diferencia de una detectaron solamente 4 patrones isoenzimáticos en sola zona de actividad encontrada para L. shannonii toda la muestra. Este sistema isoenzimático mostró por Chamberlain et al. (1996). De igual forma, se pudo constatar que no aparecieron isoformas Adh ni Mdh en el tejido foliar, lo cual está en correspondencia con lo planteado por Menezes et al. (1995), quienes informaron que en condiciones normales, la actividad enzimática de estos sistemas desaparece en etapas muy tempranas del desarrollo de las plantas, a pesar de que los mismos pueden inducirse cuando se presentan condiciones de anaerobiosis. Sin embrago, no debe descartarse además, que esto puede deberse a problemas de concentración de las muestras o de sensibilidad de la técnica. En el dendrograma (Figura 2) se observa la formación de tres grupos (I, II y III). El grupo I estuvo conformado por dos subgrupos (IA y IB). El subgrupo IA incluyó a todas las accesiones de L. leucocephala, dos accesiones de L. lanceolata (CIAT-17255 y CIAT-17501) y una de L. diversifolia (CIAT-17270), mientras que el subgrupo IB resultó heterogéneo, en el cual se incluyeron las especies L. macrophylla (3), L. esculenta (1), L. lanceolata (1) y L. diversifolia (1). Por su parte, el grupo II estuvo conformado por la accesión L. macrophylla CIAT-17232, mientras que en el grupo III se encontraron las accesiones L. macrophylla CIAT-17238 y L. esculenta CIAT-17225. De manera general, las especies L. macrophylla, L. lanceolata y principalmente, L. esculenta y L. diversifolia no formaron grupos bien definidos en el dendrograma. CONCLUSIONES Y RECOMENDACIONES El análisis de diversidad genética permitió diferenciar a la especie L. leucocephala con mayor claridad que el resto, aunque no se ganó en diferenciación dentro de la especie. Los sistemas isoenzimáticos permitieron detectar diferencias entre las accesiones, siendo las esterasas las más polimórficas. Además se comprobó una alta resolución electroforética para ambos sistemas, lo cual indica su utilidad en la evaluación del polimorfismo de Leucaena. Se recomienda realizar otras investigaciones con las accesiones seleccionadas en las que se relacione el polimorfismo isoenzimático con los marcadores moleculares y con los principales caracteres morfoagronómicos evaluados. LITERATURA CITADA
Copyright 2008 - Zootecnia Tropical The following images related to this document are available:Photo images[zt08057t1.jpg] [zt08057f1.jpg] [zt08057f2.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}