 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
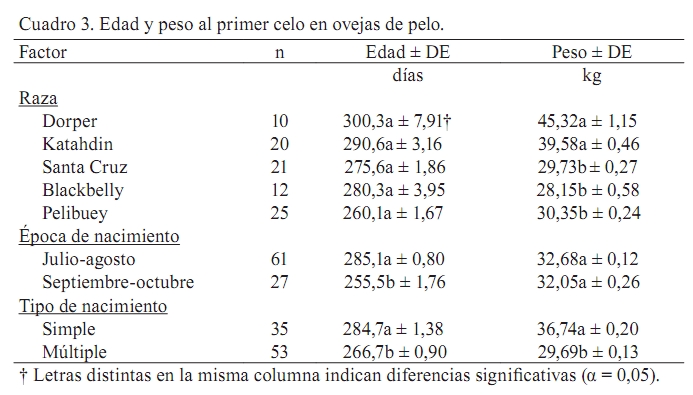
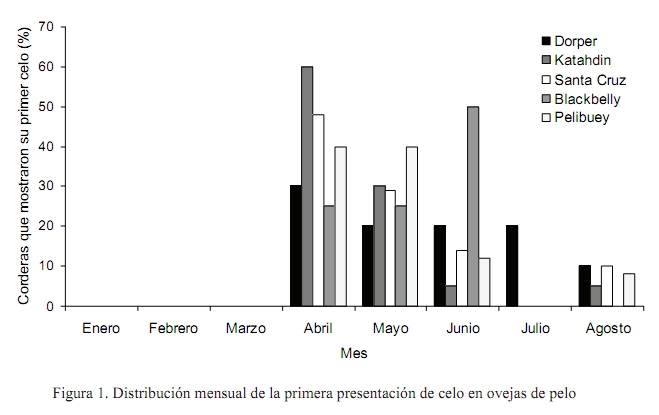
Zootecnia Tropical, Vol. 26, No. 4, 2008, pp. 465-473 Artículos Científicos Pubertad en hembras de cinco razas ovinas de pelo en condiciones de trópico seco Effect of genotype on puberty in hair sheep ewe lamb on tropical areas Rafael Zavala Elizarraraz, Jorge R. Ortiz Ortiz*, Julio P. Ramón Ugalde, Paula Montalvo Morales, Angel Sierra Vásquez y José R. Sanginés García División de Estudios de Postgrado e Investigación, Instituto Tecnológico de Conkal. Conkal, Yucatán. México. *Correo electrónico: jortiz@itconkal.edu.mx Recibido: 30/10/07 Aceptado: 12/11/08 Code number: zt08082 RESUMEN El objetivo fue estudiar el efecto del genotipo de ovejas de pelo sobre la edad y peso corporal al primer cuerpo lúteo y al primer celo observado en condiciones de trópico seco. Se utilizaron 94 corderas de las razas Pelibuey, Blackbelly, Dorper, Katahdin y Santa Cruz. Las corderas estuvieron en pastoreo rotacional 7 h/d en praderas de Cynodon nlenfluensis y se les ofreció 500 g/d de un alimento con 16% de PC. Las endoscopias se realizaron cada 28 días a partir de los cinco meses de edad monitoreando los ovarios con un endoscopio de 6,5 mm y de manera simultánea se realizó el pesaje corporal. Se utilizaron como criterios de inicio de pubertad la presencia de un cuerpo lúteo y la manifestación del primer celo. Los datos se analizaron con un modelo lineal que incluyó como variables independientes raza, época de nacimiento y tipo de nacimiento y como variables dependientes edad y peso a la pubertad, número de cuerpos lúteos y folículos. La edad y peso al primer cuerpo lúteo detectado fue mayor (P<0,05) en las ovejas Dorper (292,3 d; 43,2 kg) y Katahdin (272,6 d; 36,5 kg) en comparación con Blackbelly (250,1 d; 24,7 kg), Pelibuey (231,8 d; 27,6 kg) y Santa Cruz (252,3 d; 28,5 kg). La edad promedio al primer cuerpo lúteo para los cinco genotipos fue 252,5 d. La época de nacimiento influyó en la edad (P<0,05), pero no en el peso al primer cuerpo lúteo. Las corderas nacidas en el período julio-agosto promediaron 284,7 d, respecto a las nacidas en septiembre-octubre con 266,7 d. Las corderas de nacimiento simple tuvieron mayor edad y peso (P<0,05) (266,6 d; 34,1 kg) que aquellas de nacimiento múltiple (242,5 d; 28,6 kg). Los promedios obtenidos en la variables estudiadas resultaron superiores, pero con tendencias similares (P>0,05) cuando la pubertad se determinó con el primer celo observado, excepto el efecto de la época de nacimiento sobre el peso corporal. Los resultados evidencian mayor precocidad para las razas Pelibuey, Blackbelly y Santa Cruz que para Dorper y Katahdin. Palabras clave: pubertad, cuerpo lúteo, celo, ovejas de pelo, genotipo. ABSTRACT The objective was to evaluate the effect of hair sheep ewe lamb genotype on age and body weight to first corpus luteum and first observed estrous in dry tropic conditions. The study was performed with 94 ewe lambs Pelibuey (25), Blackbelly (12), Dorper (12), Katahdin (23), and Saint Croix (22). Ewe lambs were maintained in rotational grazing for 7 h/d on Cynodon nlemfuensis and were supplemented with 500 g/d of concentrate (16% CP). Endoscopies and lamb weight were recorded every 28 d from 150 d of age. A 6.5 mm diameter endoscope was used to determine the ovarian activity, follicular growth, luteal structures or scars present from the previous cycle. Presence of corpus luteum and first estrous behavior were used as puberty indicators. The data were analyzed with a linear model that included as independent variables breed, lambing season, and lambing type, and as dependent variables, age and weight at puberty, number of follicles, and corpus luteum. The age and weight to first corpus luteum observed were higher (P<0.05) in Dorper ewe lambs (292.3 d; 43.2 kg) and Katahdin (272.6 d; 36.5 kg) than Blackbelly (250.1 d; 24.7 kg), Pelibuey (231.8 d; 27.6 kg), and Saint Croix (252.3 d; 28.5 kg). The onset of puberty in evaluated breeds was 252.5 d of age. The season of birth affected the age (P<0.05), but not the weight of first corpus luteum observed. Ewe lambs born in July-August had 284.7 d of age, but on the other hand, those born in September-October were 266.7 d of age. Single ewe lambs were oldest and heavier (P<0.05) (266.6 d; 34.1 kg) than multiple birth ewes (242.5 d; 28.8 kg). The mean of variables studies were higher, but similar tendencies (P>0.05) were obtained with first estrous behavior, except the effect of birth seasons on live weight. These results demonstrate higher precocity in Pelibuey, Blackbelly, and Saint Croix breeds than Dorper and Katahdin. Keywords: puberty, corpus luteum , estrous behavior, hairsheep ewe lambs, genotype. INTRODUCCIÓN Los genotipos de ovinos de pelo predominantes en las regiones tropicales de México hasta mediados de la década pasada fueron los pertenecientes a las razas Pelibuey y Blackbelly (Segura et al., 1996); debido a que la ovinocultura estaba orientada de manera predominante a complementar la economía familiar de traspatio, por lo que se desarrollaba en sistemas extensivos de producción. En la actualidad se observa un crecimiento constante del sector y, en consecuencia, la producción ovina se ha intensificado, por lo que gradualmente ha habido un incremento en la introducción de razas como la Katahdin, Dorper y Santa Cruz, las cuales poseen mayor capacidad de crecimiento, sin embargo, poco se conoce de su potencial reproductivo. Considerando el alto valor del pie de cría de estas razas y la necesidad de disminuir los costos de producción en los sistemas intensivos es necesario conocer el inicio de la pubertad en los diferentes genotipos bajo las condiciones del trópico, pues de esta etapa fisiológica dependerá el inicio de la vida productiva de los reemplazos así como el número de corderos producidos, incidiendo en la productividad total de la hembra. En general, se acepta que la pubertad está influenciada principalmente por factores genéticos (Evans et al., 1991; Bunge et al., 1993; Schoeman et al., 1993; Wildeus, 1997) y por factores ambientales, tales como las condiciones climáticas (Evans et al., 1991; Schoeman et al., 1993), estado nutricional posdestete (González-Reyna et al., 1983, 1991; Albarran et al., 2001; Aguilar 2002), época de nacimiento (Cruz et al., 1983), tipo de parto y edad al destete (Fuentes et al., 1990), la presencia del macho (Rodríguez et al., 1986), entre otros. Bajo condiciones tropicales de México, en la raza Pelibuey y Blackbelly se ha documentado la pubertad (Castillo 1977; Cruz et al., 1983; Velázquez et al., 1995), pero se desconoce para corderas Dorper, Katahdin y Santa Cruz. Por lo anterior, el objetivo del presente trabajo fue determinar el efecto del genotipo de ovejas de pelo y algunos factores ambientales sobre la edad y peso a la pubertad. MATERIALES Y MÉTODOS El presente estudio se realizó en el Centro de Selección y Reproducción Ovina del Instituto Tecnológico de Conkal, Yucatán, México, el cual se ubica a 20° 59′ N y 89° 39′ O. El clima predominante es cálido subhúmedo con lluvias en verano (Aw0), con 900 mm de precipitación anual, altura de 9 msnm y una temperatura media anual de 26,5°C (García, 1981). Se utilizaron 94 corderas puras de las razas Dorper (n = 12), Katahdin (n = 23), Santa Cruz (n = 22), Blackbelly (n = 12) y Pelibuey (n = 25), nacidas entre los meses de Julio y Octubre. En el Cuadro 1 se presenta la distribución de las corderas por raza, tipo y época de nacimiento. El manejo de las madres, las corderas y los sementales consistió en pastoreo rotacional de 7 h diarias, en potreros de pasto estrella de África (Cynodon nlemfuensis). Al regreso del pastoreo, en corrales de encierro, las hembras en gestación fueron suplementadas con 150–200 g de un alimento con base en sorgo y pasta de soya (16% PC), en tanto las madres en lactancia y los machos recibieron 400 y 300 g animal-1 d-1, respectivamente. Las corderas permanecieron con sus madres desde el nacimiento hasta el destete a los 90 d de edad y a partir de la sexta semana se les proporcionó entre 100 a 150 g de alimento con base en sorgo y pasta de soya (18% PC). Al destete todas las corderas se desparasitaron con ivermectina (Cidectin® NF- 1%) y vacunaron contra Pasterella hemolitica y clostridiasis. A partir de ese momento las corderas permanecieron bajo las mismas condiciones de manejo y alimentación al resto del rebaño pero suplementadas con 500 g animal-1 d-1 de un alimento comercial con 15% de PC, a base de sorgo molido y entero, pasta de canola y subproducto de ajonjolí. La pubertad se determinó a través de dos métodos: por endoscopías (detección del primer cuerpo lúteo) y por detección directa del estro (manifestación clínica de estro con la conducta sexual correspondiente). Las endoscopías se efectuaron de acuerdo a la metodología propuesta por Oldham y Lindsay (1980). Las cirugías se realizaron a partir de que las corderas cumplían los cinco meses de edad, empleando un endoscopio Karl Storz de 6,5 mm de diámetro. Las endoscopías se ejecutaron cada 28 d; en cada una de ellas, los folículos detectados se contaron y ordenaron en tres clases: menor a 1, de 2 a 4 y mayor o igual a 5 mm de diámetro. En las corderas que presentaron un cuerpo lúteo (CL) y/o cuerpo blanco del ciclo anterior no se volvió a realizar otra endoscopia, siendo considerado este hecho como el inicio de la pubertad. La detección del celo se inició a partir de los cinco meses de edad con tres machos vasectomizados, los cuales se rotaban por intervalos de una semana. Los machos se introducían al corral de encierro a las 7:00 y 17:00 h por periodos de una h. La manifestación de signos evidentes de celo por primera vez en las corderas se consideró como el segundo criterio de inicio de pubertad. Los pesos corporales se registraron momentos previos a las endoscopias y al momento de detectarse el celo. Los datos se sometieron al análisis de varianza para modelos lineales (PROC GLM de SAS, 2000) y para la comparación de medias se utilizó la prueba de Tukey. Para las variables dependientes edad y peso a la pubertad a través del primer cuerpo lúteo (CL) y primer celo observado, se utilizó el siguiente modelo estadístico: уijkl = µ + αi + βj + αβij + θk + α θik + βθjk + Єijkl donde: уijk = variable de respuesta en la l-ésima repetición del i-ésimo tipo racial en la j-ésima época de nacimiento en el k-ésimo tipo de nacimiento, En el modelo no se consideró la triple interacción entre raza, época de nacimiento y tipo de nacimiento, debido a que es un modelo desbalanceado y al momento de realizar el análisis se encontraron sumas de cuadrados no estimados por la existencia de celdas vacías, es decir, sin observaciones. Para las variables número de folículos y cuerpos lúteos así como el tamaño (mm) de los folículos se consideraron los valores promedios ponderados por raza de las siete endoscopías realizadas mensualmente a lo largo del período experimental. El análisis fue con el siguiente modelo estadístico: уij = µ + αi + Єij donde: yij = variable de respuesta en la i-ésima raza y la j-ésima observación, En el modelo no se consideraron los factores de variación de época y tipo de nacimiento y las diferentes interacciones, ya que al hacerlo se hubiesen generado 20 subclases por cada endoscopía y durante el periodo experimental se realizaron siete endoscopias, por lo que no hubo la suficiente cantidad de observaciones. En consecuencia se analizaron los promedios ponderados para cada raza durante las siete endoscopías, haciendo énfasis en el efecto de raza. La comparación de medias se realizó mediante la prueba de Tukey a un α = 0,05. Así mismo, se graficó la frecuencia de celos detectados. RESULTADOS Y DISCUSIÓN Durante el desarrollo del experimento, murió una cordera de la raza Katahdin y se presentaron adherencias en dos corderas de raza Pelibuey, lo que impidió observar la presencia de algún cuerpo lúteo. Por otra parte, cinco corderas (dos de raza Dorper, dos Katahdin y una Santa Cruz) no mostraron presencia de cuerpo lúteo o cicatrizal, por lo que no alcanzaron la pubertad. No se pudo precisar si existió algún efecto de las lesiones ocasionadas por las laparoscopias sobre la expresión del celo en algunas corderas. Raza No hubo evidencia de interacciones (P>0,05) entre la raza y el tipo y época de nacimiento y entre el tipo y época de nacimiento, por lo que solamente se presentan los resultados de los efectos principales. El análisis mostró diferencias (P<0,05) entre razas, para la edad y peso al primer CL (Cuadro 2). Las corderas de la razas Dorper y Katahdin, presentaron un primer CL observado 59 y 41 d más tarde que la raza Pelibuey, Así mismo, tuvieron 15,6 y 8,9 kg más de peso corporal con respecto a las corderas de raza Pelibuey. La diferencia para las corderas Santa Cruz, respecto a las Pelibuey en las mismas variables fue de 21 d y 0,95 kg. Por otra parte, cuando la pubertad se determinó al primer celo observado (Cuadro 3), solamente se encontraron diferencias para el peso corporal entre razas (P<0,01). Las razas Dorper y Katahdin pesaron al menos 15 y 9 kg más con respecto a las razas Santa Cruz, Pelibuey y Blackbelly. Las cinco razas, para la edad al primer celo observado, manifestaron la misma tendencia que la pubertad al primer CL, es decir las razas sintéticas tuvieron promedios de edades más altas. El efecto de la raza sobre la edad y peso a la pubertad ha sido demostrado por otros autores en estudios comparativos en condiciones de clima templado, tanto en ovejas de lana y de pelo (Evans et al., 1991; Pappa-Michailidou et al., 1999; Papachristoforou et al., 2000), como entre sus cruzas (Lassoued y Rekik, 2001) y en cruzas con ovinos de pelo (Bunge et al., 1993). En condiciones de trópico, la mayoría de los estudios proporcionan valores de edad y peso a la pubertad en la raza Pelibuey en el rango de 245 a 326 d y de 18,4 a 25 kg (Castillo, 1977; González-Reyna et al., 1983; Valencia y González, 1983; Fuentes et al., 1990; González-Reyna et al., 1991), valores que no coinciden en la edad ni en el peso a la pubertad obtenidos para el Pelibuey en el presente estudio. A su vez, las corderas Blackbelly, considerada como la única raza prolífica tropical (Fahmy y Mason, 1996), alcanzaron la precocidad en un rango que varia de 215 a 407 d con un peso de 25 a 26,3 kg (Bradford et al., 1983; Velázquez 1994; Rastogui 1996). La edad al primer CL (252 d) estuvo en el rango reportado para Santa Cruz en trópico (180 a 270 d), pero no para clima templado (150-180 d) donde dicha raza exhibe mayor precocidad (Evans et al., 1991). Cloete et al. (2000), en una revisión bibliográfica sobre la raza Dorper citan un rango de valores para edad y peso a la pubertad de 213 a 245 d y de 39 a 50.8 kg en ambientes templados y semiáridas, valores que coinciden para el peso obtenido en este estudio (43 a 45 kg), pero no para la edad (291 a 300 d). Resultados de Schoeman et al. (1993) indican que la raza Dorper es más pesada a la pubertad y tiene menor tasa de ovulación que Romanov y sus cruzas con Romanov y Finnsheep, evidenciando que es una raza de madurez tardía, conclusión que apoya la tendencia de nuestras observaciones. Similarmente, para la raza Katahdin no hay reportes en clima tropical; sin embargo, Fahmy y Mason (1996) para condiciones templadas observaron ovejas precoces a los seis meses de edad con un peso de 50 a 75 kg, los cuales no coinciden a los de este estudio. Las diferencias en pubertad entre razas pueden ser debidas a su estructura genética que influye en la capacidad de adaptación al ambiente y a las condiciones impuestas tanto por el clima como por los sistemas de manejo. Las cinco razas estudiadas son de pelo, pero sólo se consideran razas puras Pelibuey y Blackbelly, en tanto las restantes son consideradas sintéticas. En general, los resultados de este estudio, obtenidos en clima tropical seco en un sistema semintensivo, evidenciaron que los genotipos sintéticos demoraron 27 d más en manifestar el primer CL y 20 d más en exhibir conducta sexual que las razas puras; así mismo los valores respectivos para el peso fueron de 8,2 y 7 kg más, respectivamente. Estos resultados sugieren revisar las políticas de reemplazos del pie de cría y su manejo reproductivo, pues es posible que las diferencias se magnifiquen o disminuyan según se trate de sistemas extensivos o intensivos. Época y tipo de nacimiento La época de nacimiento afectó la edad (P<0,01), pero no el peso a la pubertad en las corderas de este estudio (Cuadro 2 y Cuadro 3). Tanto en la edad al primer CL como al primer estro, las corderas nacidas en verano tuvieron 30 d más de edad que aquellas nacidas en el otoño. Asimismo, el tipo de nacimiento influyo tanto en la edad como en el peso a la pubertad (P<0,05) (Cuadro 2 y Cuadro 3). Las corderas procedentes de nacimiento simple requirieron de 19 a 29 d y de 5 a 7 kg de peso más para el inicio de la actividad sexual que las de parto doble. En este resultado probablemente influyó el número de partos simples de las razas Dorper y Katahdin que representaron 56% de las corderas. Por el contrario, solamente 18% de las corderas Dorper y Katahdin pertenecieron al grupo de nacimiento doble. Nuestros resultados coinciden con la literatura debido a que existe una marcada influencia de la estación de nacimiento y tipo de nacimiento en la pubertad. Corderas nacidas en primavera-verano (mayo a julio) demoran mas (88 d) en su edad al primer parto, por lo que manifiestan la pubertad a mayor edad que las nacidas en las otras épocas del año (Cruz et al., 1983). Resultados con tendencias similares fueron encontrados por Ponce de León et al. (1981) y han sido confirmados por González-Stagnaro (2002). La demora en el inicio de la pubertad y edad al primer parto para las nacidas en primavera-verano se asocia a una disminución en las calidad y cantidad de las fuentes de energía (usualmente pastos y forrajes) de dicha época, que influyen en el desarrollo corporal de las corderas que han sido destetadas previo a su inicio. Un aspecto que contribuye a explicar los efectos de la época de nacimiento y tipo de parto es el estado nutricional pre y posdestete, pues la manipulación de la alimentación puede adelantar o retrasar el inicio de la pubertad (Valencia y González, 1983; González-Reyna, 1991). Niveles crecientes de suplementación del 0, 1, 2 y 3% en corderas Pelibuey acortan el inicio de la pubertad de 329 a 261 d; sin embargo, no producen diferencias en el peso corporal (28,9 a 26,5 kg) (Velázquez et al., 1995). A su vez, González-Stagnaro (2002) demostró que un déficit nutricional retrasa el desarrollo del tracto genital, que presenta un crecimiento alométrico entre los 230 a 310 días de edad, periodo en el que el tracto genital creció mas en longitud y volumen respecto a su propio peso, y por ende, explica la influencia de la alimentación en la edad a la pubertad. La suplementación a hembras adultas durante la gestación no influye en el peso ni la edad a la pubertad, cuando se utiliza como indicador la primera manifestación de celo (21,7 kg y 238 d vs. 21,8 kg y 224,6 d) expresado por sus hijas (Albarran et al., 2001). Estructuras ováricas En el Cuadro 4 se presentan los promedios ponderados de las estructuras ováricas durante las siete endoscopias, en donde se observaron diferencias (P<0,05) entre razas para el número de folículos, en tanto que en el tamaño de los folículos y el número de CL fueron similares (P>0,05). El mayor número de folículos se observó en las corderas Katahdin, Blackbelly, Pelibuey y Dorper y el menor número en las corderas Santa Cruz. Por otra parte, el mayor diámetro folicular se encontró en las corderas Dorper y Blackbelly. Las diferencias encontradas en el número de folículos y no observadas en los cuerpos lúteos permiten plantear la posibilidad de atribuirlo a una mayor atresia folicular en dichas razas (con excepción de la raza Santa Cruz), quedando pocos folículos reclutados y seleccionados para el crecimiento, maduración y ovulación, no obstante esta hipótesis debe probarse en futuros ensayos. Para razas ovinas prolíficas de clima templado en hembras adultas, se han encontrado tasas ovulatorias de 1,87 para Finnsheep y 2,21 en Merino Booroola (Young y Dickerson, 1991). Similarmente, pero en corderas F1 provenientes de padres Finnsheep, Combo 6, Booroola Merino, Saint Croix y Barbados los números de cuerpos lúteos observados fueron 2,41 1,69 2,89 1,69 y 1,59, respectivamente (Bunge et al., 1993). Barroso (1995) encontró una tasa ovulatoria en clima tropical para hembras multíparas de raza pura Blackbelly en empadre de invierno de 2,75 a 2,27 según recibieran o no suplementación energética. En todos los casos los valores resultaron superiores a los del presente estudio. De este modo, es probable que operen diferentes sistemas de control para generar una elevada tasa de ovulación en algunas razas. Celos A pesar de que la observación en las conductas de celo se inició a partir del mes de enero, las primeras manifestaciones se registraron en abril, en 40,9% de las corderas, el acumulado para mayo fue 68.8% y el restante 25,8% entraron en celo entre junio y agosto. Solo 5,4% de las corderas no mostraron celo durante el periodo que duró el trabajo. En la Figura 1 se presenta la distribución mensual del primer celo en las corderas de los diferentes tipos raciales. Se ha observado que las ovejas Pelibuey disminuyen la actividad ovárica en los meses de enero a abril y se relaciona con la disponibilidad de forraje (González-Reyna et al., 1991; Higuera et al., 2000). En el año en que se realizó el presente trabajo, las lluvias de verano se adelantaron y ocurrieron a finales de marzo, aumentando la disponibilidad y calidad del forraje, lo que pudo haber favorecido la presentación de estros durante los meses de abril y mayo. CONCLUSIONES El genotipo y el tipo de nacimiento influyeron en la edad y el peso a la pubertad, siendo las ovejas locales Pelibuey y Blackbelly más precoces que Dorper y Katahdin, siendo además las corderas de nacimiento simple, de mayor edad y peso a la pubertad que las múltiples. AGRADECIMIENTOS El presente trabajo es parte de la tesis de Maestría en Ciencias del primer autor, el cual fue posible gracias al apoyo financiero de la Dirección General de Educación Tecnológica Agropecuaria (DGETA). LITERATURA CITADA
Copyright © 2008 - Zootecnia Tropical The following images related to this document are available:Photo images[zt08082c4.jpg] [zt08082c3.jpg] [zt08082c1.jpg] [zt08082f1.jpg] [zt08082c2.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}