 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
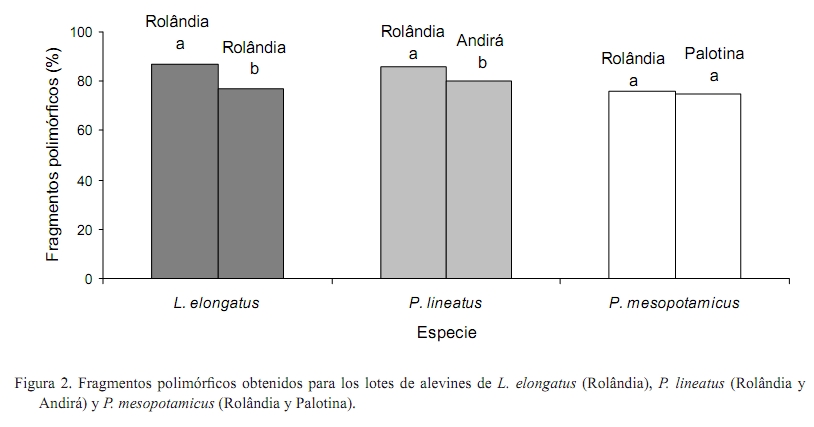
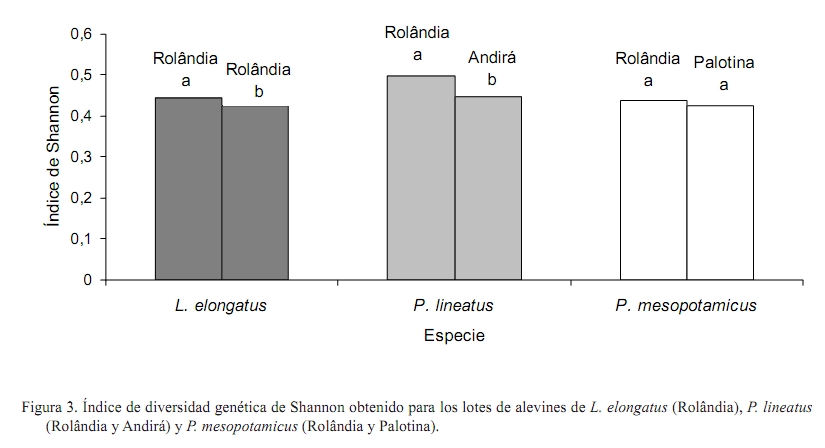
Zootecnia Tropical, Vol. 26, No. 4, 2008, pp. 515-522. Artículos Científicos Caracterización genética de lotes de peces usados en programas de repoblamiento y su importancia en la conservación genética en la piscicultura Genetic characterization of fish stocks used in stocking programas and their importance in the fish farm genetic conservation Nelson M. Lopera Barrero1*, Ricardo Pereira Ribeiro1, Jayme A. Povh2, Patrícia C. Gomes1, Lauro Vargas1 y Sheila Nogueira de Oliveira1 1 Universidade Estadual de Maringá, Grupo de Pesquisa PeixeGen, Centro de Ciências Agrárias, Av. Colombo, 5790, Bloco J45, CEP 87020-900, Maringá, PR, Brasil. *Correo electrónico: nelson.peixegen@gmail.com. Recibido: 13/1107 Aceptado: 27/06/08 Code number: zt08087 RESUMEN En las últimas décadas se ha verificado la desaparición de varias especies de peces debido principalmente a impactos generados por acciones humanas. Programas de repoblamiento vienen siendo cada vez más usados como métodos de conservación de la ictiofauna. Sin embargo, sin una correcta orientación genética y reproductiva de los lotes utilizados en estos programas, poblaciones naturales de peces y el ecosistema pueden ser afectados. El objetivo del siguiente estudio fue determinar la variabilidad genética de seis lotes de peces usados en programas de repoblamiento, mediante el marcador molecular RAPD. Se analizaron 180 alevines de tres especies de peces (Leporinus elongatus, Piaractus mesopotamicus y Prochilodus lineatus) en tres estaciones piscícolas, ubicadas en las ciudades de Rolândia, Andirá y Palotina en el estado de Paraná, Brasil. Los valores de variabilidad genética estimados por el porcentaje de fragmentos polimórficos y por el índice de diversidad de Shannon mostraron una alta variabilidad genética entre los lotes de L. elongatus y P. lineatus, debido posiblemente al efecto fundador y al manejo reproductivo adoptado en cada piscícola. Se determinó que existió baja diferenciación genética entre los lotes de P. mesopotamicus. Los resultados de este estudio posibilitaran el correcto manejo reproductivo y genético de los lotes de cada piscícola y la orientación objetiva de programas de repoblamiento, permitiendo la conservación de la variabilidad genética, factor de gran importancia en ambientes en cautiverio. Palabras clave: Leporinus elongatus, Piaractus mesopotamicus, Prochilodus lineatus, repoblamiento, variabilidad genética. ABSTRACT In the last decades it has been verified the decrease and extinction of fish several species mainly to impacts generated by human actions. Stocking programs are being used as conservation methods of the ichthyofauna. However, without a correct genetic and reproductive orientation of the stocks used in these programs, natural fish populations and the ecosystem can be affected. The objective of the following study was to determine the genetic variability of six fish stocks used in stocking programs, by means of the RAPD molecular marker. There were analyzed 180 juveniles of three fish species (Prochilodus lineatus, Piaractus mesopotamicus, and Leporinus elongatus) from three fish farms, located in the Rolândia, Andirá, and Palotina cities in Paraná state, Brazil. The genetic variability values estimated by the percentage of polymorphic fragments and by the Shannon diversity index showed a high genetic variability between the P. lineatus and L. elongatus stocks, due possibly to the founder effect and the reproductive management adopted in each fish farm. It was determined that low genetic differentiation existed among the P. mesopotamicus stocks. The results of this study facilitated the correct reproductive and genetic management of the stocks of each fish farm and the objective orientation of stocking programs, allowing the conservation of the genetic variability, factor of great importance in captivity environments. Keywords: genetic variability, Leporinus elongatus, Piaractus mesopotamicus, Prochilodus lineatus, stocking. INTRODUCCIÓN La deforestación y su consecuente reducción de las fuentes de alimento, la construcción de hidroeléctricas y drenajes para el área agrícola (Hatanaka et al., 2006), la degradación de la calidad del agua en función de la contaminación (Hori et al., 2006), el ecoturismo mal planeado (Sabino y Andrade, 2003) y la falta de conocimiento taxonómico (Agostinho et al., 2005) han llevado a la disminución y extinción de varias especies de peces en los últimos años. Entre esas especies, Leporinus elongatus, Piaractus mesopotamicus y Prochilodus lineatus, peces migratorios nativos brasileños conocidos regionalmente como piapara, pacu y curimba (Furuya, 2001), vienen presentando una reducción progresiva de sus poblaciones naturales en consecuencia de las mudanzas ambientales generadas principalmente por acciones humanas. De las diversas herramientas empleadas para reducir esos impactos sobre las poblaciones de peces, la practica de repoblamiento de los ríos viene tornándose cada vez más común (Hilsdorf et al., 2006; Agostinho y Gomes, 2006). Sin embargo, sin un apoyo científico que permita su correcta orientación, estos programas pueden tornarse en una amenaza mayor para los ecosistemas y para las poblaciones naturales de peces (Agostinho et al., 2005). Los análisis genéticos de lotes de pisciculturas representan informaciones de gran importancia para conseguir resultados expresivos en la producción y en la conservación de peces (Lopera Barrero, 2007), ya que la perdida de variabilidad genética en lotes de estaciones piscícolas debido al inadecuado manejo reproductivo (Frost et al., 2006) o por deficiencias en el número efectivo de reproductores (Aho et al., 2006) puede producir problemas de endogamia, adaptabilidad y supervivencia de progenies usadas en programas de repoblamiento (Povh et al., 2008). Esos problemas pueden consecuentemente afectar las poblaciones naturales de peces (Sønstebø et al., 2007) y el ecosistema en general, pudiendo conducir la especie a la extinción (Agostinho et al., 2005). Por tanto, el objetivo de este estudio fue analizar la variabilidad genética de seis lotes de peces usados en programas de repoblamiento, mediante el marcador molecular RAPD. Los resultados permitirán orientar objetivamente estos programas realizados en ríos brasileños y así evitar la perdida de variabilidad genética en los lotes de reproductores, en las progenies usadas en programas de repoblamiento y en las poblaciones naturales. MATERIALES Y MÉTODOS Material biológico Muestras de aleta caudal (30 de cada lote) fueron recolectadas en seis lotes de alevinos de piscícolas ubicadas en las ciudades de Rolândia (dos lotes de L. elongatus y uno de P. mesopotamicus y P. lineatus), Palotina (un lote de P. mesopotamicus) y Andirá (un lote de P. lineatus), en el estado del Paraná, Brasil, los cuales son usados en programas de repoblamiento de los ríos Paraná y Paranapanema (Figura 1). Extracción de ADN Para la extracción de ADN se utilizó la metodología descrita por Lopera-Barrero et al. (2008). En micro-tubos conteniendo las aletas, se adicionaron 550 µL de tampón de lisis (50 mM Tris-HCl, 50 mM EDTA, 100 mM NaCl y 1% SDS) y 7 µL de proteinasa K (200 µg/mL). Se incubaron en baño-maría a 50ºC por 12 h. El ADN se lavó con 600 µL de solución de NaCl (5 M) y se centrifugó por 10 min a 12.000 rpm. El sobrenadante conteniendo el ADN se transfirió para nuevos micro-tubos, se precipitó con 700 µL de alcohol etílico absoluto e se incubó por 1 h a -20ºC. El ADN se centrifugó, se lavó con 700 µL de alcohol etílico 70%, se suspendió en 80 µL de tampón TE (10 mM Tris pH 8,0 y 1 mM EDTA) y se trató con 7 µL de RNAsa (30 µg/mL) en baño-maría a 37ºC por 1 h. Se guardó en el congelador a -20ºC. Cuantificación y amplificación del ADN El ADN se cuantificó en espectrofotómetro Shimadzu (UV 1601, EE.UU.) con amplitud de onda de 260 nm. Las muestras se diluyeron para una concentración de 10 ng/µL. Para confirmar la calidad del ADN, se realizó una electroforesis en gel de agarosa 1%, conducida en tampón TBE 1X (500 mM Tris-HC1, 60 mM ácido bórico y 83 mM EDTA) por 1 h a 70 V. Las condiciones de amplificación se basaron en los procedimientos descritos por Williams et al. (1990), con modificaciones. El ADN genómico se amplificó en un volumen de reacción de 15 µL, en el cual se utilizó tampón 1X Tris-KCl, 2,5 mM de MgCl2, 0,46 µM de iniciador, 0,2 mM de cada dNTP, una unidad de Platinun Taq ADN Polimerasa (Invitrogen®, EE.UU.) y 10 ng de ADN. El ADN se desnaturó a 94ºC por 4 min y enseguida se realizaron 40 ciclos, cada uno consistiendo de 1 min de desnaturación a 94ºC, 90 seg de anelamiento del iniciador a 40ºC y 2 min para extensión a 72ºC. Despues se realizó una extensión final a 72ºC por 5 min. Las reacciones de RAPD se amplificaron en un termociclador (Eppendorf Mastercycler Gradient, EE.UU.). Se evaluó la amplificación de 60 diferentes iniciadores de 10 bases de los kits OPA OPX y OPW (Operon Technologies Ltd., EE.UU.) siendo escogidos los que presentaron mejor definición y reproductibilidad. Los productos de amplificación se separaron en gel de agarosa 1,5%. Se utilizaron 15 µL del producto amplificado y 2 µL de tampón de muestra (40% sacarosa y 0,25% azul de bromofenol) en electroforesis horizontal. La electroforesis se condujo en tampón TBE 0,5X (45 mM Tris-Borato y 1 mM EDTA) por 4 h a 70 V. Los geles de cuantificación y amplificación se visualizaron sobre radiación UV, despues de su exposición con bromuro de etídio (0,5 µg/mL) por 1 h. La imagen se fotografió utilizando el programa Kodak EDAS (Kodak 1D Image Analysis 3.5, EE.UU.). Análisis estadístico El tamaño de los fragmentos obtenidos con las amplificaciones se estimó por comparación con el padrón ladder 100 pb (Invitrogen®, EE.UU). La presencia o ausencia de fragmentos de tamaños moleculares idénticos se usó para la construcción de una matriz con base en el cálculo del coeficiente de similitud de Jaccard, codificando 1 como presencia de fragmento y 0 como su ausencia. La variabilidad genética se determinó por el porcentaje de fragmentos polimórficos y por el índice de diversidad genética de Shannon a través del programa PopGene 1.31 (Yeh et al., 1999). RESULTADOS Y DISCUSIÓN Los ocho iniciadores seleccionados OPA16 (5’-AGCCAGCGAA-3’), OPW01 (5’-CTCAGTGTCC-3’), OPW02 (5’-ACCCCGCCAA-3’), OPW03 (5’-GTCCGGAGTG-3’), OPW08 (5’-GACTGCCTCT-3’), OPW19 (5’-CAAAGCGCTC-3’), OPX01 (5’-CTGGGCACGA-3’) y OPX03 (5’-TGGCGCAGTG-3’) produjeron un total de 70 fragmentos, con tamaños entre 200 a 1200 pb. La variabilidad genética, estimada por el valor de fragmentos polimórficos (FP) y por el índice de diversidad genética de Shannon (IS), fue alta entre los lotes de L. elongatus y de P. lineatus. Por otro lado, fue encontrada baja diferenciación genética entre los lotes de P. mesopotamicus de las ciudades de Rolândia y Palotina (Figura 2 y Figura 3). Según Moreira et al. (2007), una perdida de variabilidad genética en la piscicultura siempre es esperada cuando existe un mal manejo reproductivo debido al cruzamiento de individuos emparentados, lo que consecuentemente aumenta el coeficiente de endogamia (Kang et al., 2006) y reducirá el número efectivo de reproductores (Frost et al., 2006). Esta situación es bastante común en piscícolas, ya que el método mas ampliamente usado en la formación de nuevos lotes es la selección de individuos con características visuales favorables (de mayor tamaño y con mejores condiciones reproductivas), involucrando de esa forma el efecto boca de botella (bottleneck effect) donde debido a esa selección intencional pueden presentarse problemas de endogamia por el alto relacionamiento genético entre los individuos, ocasionando perdida de la variabilidad genética en los reproductores (Aho et al., 2006) y en sus progenies. En nuestros resultados observamos que este efecto no influenció los lotes de L. elongatus a pesar de pertenecer a la misma piscícola, siendo encontrados altos valores de variabilidad genética entre ellos (%FP = 86,96 y 76,74; IS = 0,444 y 0,425). Esto demuestra que hubo un buen manejo reproductivo, donde la formación de lotes con suficiente variabilidad genética (efecto fundador), la introducción de reproductores genéticamente divergentes y de diferentes orígenes (Freitas y Galetti Jr, 2005) y la utilización de sistemas reproductivos y de cruzamientos eficientes (Lopera Barrero, 2007; Povh, 2007) posiblemente permitió mantener el pool genético de lotes mantenidos en ambientes controlados y así poder usarlos correctamente con fines de repoblamiento. Igualmente, al analizar los lotes de P. lineatus de las piscícolas de las ciudades de Rolândia y Andirá se observó que hubo una alta diferenciación genética entre ellos (%FP=85,71 y 80; IS=0,498 y 0,448). Este resultado puede ser atribuido al efecto fundador, ya que las piscícolas fueron formadas a partir de poblaciones naturales de peces recolectadas en el rio Paraná y Paranapanema, respectivamente, existiendo una diferenciación geográfica y de manejo. Gomes (2007) al analizar tres lotes de L. elongatus usados en programas de repoblamiento, encontró igualmente una alta variabilidad genética entre ellos debida principalmente al efecto fundador y al eficiente manejo reproductivo de los lotes. Por otro lado, al analizar los lotes de P. mesopotamicus de las piscícolas de Rolândia y Palotina no fue verificada una alta variabilidad genética entre ellos (%FP=76,12 y 75; IS=0,438 y 0,425). Caso contrario al observado en los lotes de P. lineatus, los lotes de P. mesopotamicus fueron formados a partir de poblaciones naturales de peces recolectados en diferentes localidades del rio Paraná, siendo posible que exista una similitud genética entre ellos por la existencia de flujo génico. Esta situación es común en poblaciones naturales, como fue verificado por Leuzzi et al. (2004), los cuales al analizar poblaciones de Astyanax altiparanae de dos reservorios del río Paranapanema encontraron que hubo flujo génico entre poblaciones separadas geográficamente. Entre tanto, no es posible hacer inferencias al respecto de la validez de esta teoría ya que no hay registros sobre el lugar exacto donde los reproductores fueron capturados. Otra explicación a estos resultados puede ser la perdida de variabilidad genética que para algunos autores es normalmente irreversible (Wasko et al., 2004; Sekino et al., 2004), lo que consecuentemente aumentó la similitud genética entre los individuos de los dos lotes. Los resultados encontrados en este estudio son de gran importancia, ya que a partir de ellos el manejo reproductivo de esos lotes utilizados en programas de repoblamiento pueden ser orientados correctamente y con mayor objetividad. Las especies de importancia comercial y especialmente aquellas amenazadas de extinción como es el caso de las analizadas en este estudio, requieren un constante monitoreo genético de sus lotes mantenidos en las piscícolas y de sus poblaciones naturales. La disminución de la variabilidad genética puede tornar un programa de repoblamiento ineficiente (baja supervivencia de los juveniles en el ambiente acuático) y proporcionar impactos genéticos irreversibles en las poblaciones nativas (Povh et al., 2008) que pueden llegar a la extinción de especies. Según Sønstebø et al. (2007) y Melo et al. (2006), el cruzamiento de individuos genéticamente distintos a aquellos encontrados en una población natural puede promover la perdida de genes importantes de adaptabilidad al ambiente, que puede influenciar en la supervivencia de progenies en el ambiente natural. A partir de estas evidencias y de los resultados de variabilidad genética, podemos sugerir que el manejo reproductivo, genético y de mejoramiento de los lotes de P. mesopotamicus de la región de Rolândia y Palotina deben ser realizados de manera homogénea, como unidades genéticamente iguales y no como lotes separados. Así mismo, para aumentar la variabilidad genética de esos lotes, es necesaria la introducción de nuevo material genético (reproductores), los cuales pueden ser capturados a partir de poblaciones naturales genéticamente diferentes de los ríos Paraná o Paranapanema o también con el intercambio de reproductores entre otras estaciones piscícolas, pudiendo contribuir con un pool genético que permita la preservación de la variabilidad genética. Es recomendable realizar esta introducción e intercambio de reproductores siempre basado en análisis genéticos, ya que puede ocurrir depresión exogámica cuando los individuos son muy diferentes. De esta forma, la correcta selección de los reproductores que serán usados en la formación del lote de una piscícola y el monitoreo genético de estos pueden ofrecer bases importantes para formular estrategias de manejo reproductivo (Sønstebø et al., 2007), las cuales permitirán un intercambio de reproductores entre las estaciones piscícolas y de esa forma fragmentar ciclos de endogamia que son comunes en ambientes controlados (Moreira et al., 2007) y que pueden definir la conservación de una especie y su futuro potencial biológico (Melo et al., 2006). Por esta razón, la primera providencia a ser tomada en la implantación de una piscícola o de programas de repoblamiento, como es el caso para los lotes de L. elongatus, P. lineatus y P. mesopotamicus analizados en este estudio, es verificar la diversidad genética de ellos, de las progenies y de las poblaciones naturales en todos los periodos del año, evitando la perdida de variabilidad genética y de adaptabilidad en los lotes y poblaciones naturales de peces y al mismo tiempo verificando la eficacia de este tipo de programa de conservación y sus posibles efectos adversos en la ictiofauna y en el ecosistema. Con ese objetivo, se recomienda la participación multidisciplinar junto con la participación de la sociedad, que permita direccionar científicamente la conservación (Lopera-Barrero et al., 2007). De la misma forma, Sirol y Britto (2006) enfatizan también que la definición de cuales especies deben ser repobladas debe basarse en su importancia socio-cultural en el ambiente y principalmente en la capacidad de formar una población sustentable, que no afecte el ciclo de vida de otras especies y la biodiversidad presente en el ecosistema. Entre tanto, además del monitoreo de la variabilidad genética y de la estructura genética de todas las poblaciones de peces presentes en condiciones de cautiverio y en el ambiente natural, factores como la preservación de los bosques, la protección de los ríos, el control de la pesca y la participación de la sociedad y de las entidades publicas y privadas deben ser considerados para alcanzar la conservación del ecosistema y el éxito de los programas de repoblamiento (Lopera-Barrero et al., 2007). Con los resultados de este estudio fue posible obtener un perfil de los lotes de alevinos de estas piscícolas, su caracterización genética y la objetiva orientación genética y reproductiva que permitirá la correcta conservación de la variabilidad genética del L. elongatus, P. lineatus y P. mesopotamicus. Para eso, el marcador molecular RAPD se mostró eficaz y ofreció buenos resultados. CONCLUSIONES Los valores de variabilidad genética mostraron una alta variabilidad genética entre los lotes de L. elongatus y P. lineatus, debido posiblemente al efecto fundador y al manejo reproductivo adoptado en cada piscícola. Se determinó igualmente que existió baja diferenciación genética entre los lotes de P. mesopotamicus. Los resultados permitirán orientar objetivamente estos lotes de peces utilizados en programas de repoblamiento y así evitar la pérdida de variabilidad genética en los reproductores y en las progenies. LITERATURA CITADA
Copyright © 2008 - Zootecnia Tropical The following images related to this document are available:Photo images[zt08087f2.jpg] [zt08087f3.jpg] [zt08087f1.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}