 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Zootecnia Tropical, Vol. 27, No. 1, 2009, pp. 7-15. Artículos Científicos Evaluación de la fertilidad y eclosión en la codorniz japonesa Evaluation of the fertility and hatchability in Japanese quails Rafael Galíndez*, Vasco De Basilio, Gonzalo Martínez, Daniel Vargas, Edwin Uztariz y Patricia Mejía Universidad Central de Venezuela. Facultad de Agronomía. Instituto de Producción Animal. Apartado Postal 4579. Maracay, Aragua. Venezuela. *Correo electrónico: galindez70@yahoo.com Recibido: 22/11/2008 Aceptado: 10/02/2009 Code number: zt09002 RESUMEN Con el objetivo de estudiar el efecto de algunos factores no genéticos sobre la fertilidad y eclosión de huevos de codornices (Coturnix coturnix japonica) sometidas a diferentes modalidades de apareamiento, se realizaron tres análisis de varianza asumiendo una distribución binomial de los datos. Se utilizaron 2.613 registros para fertilidad (FER), 2.178 para eclosión de huevos totales (EHT) y 1.403 para eclosión de huevos fértiles (EHF) provenientes de codornices experimentales. Se incluyeron los efectos mes de postura, generación y tipo de apareamiento para FER y para EHT y EHF se adicionaron el peso del huevo, color y brillo de la cáscara. Se ofreció agua y alimento ad libitum. La relación macho: hembra fue 1:3. El sistema de reproducción comprendió dos modalidades: rotativo y continuo. Se encontraron promedios de 78,7 25,8 y 42,3% para FER, EHT y EHF, respectivamente. El mes de mayor (P<0,01) fertilidad fue marzo (91,1%), mientras que el menor mes fue noviembre (51,7%). La mayor eclosión (P<0,01) ocurrió en agosto (42,6 y 65,7%) y la menor se observó en abril (12,5 y 17,9%) para EHT y EHF, respectivamente. Hubo mayor fertilidad (P<0,01 86,2%) en los huevos provenientes de apareamientos continuos. Los huevos que presentaron mayor porcentaje de eclosión (P<0,01) fueron aquellos de cáscara brillante (43,9%), color oscuro (48,0%) y pesos intermedios (10,1 a 11,0 g; 44,0%). Se concluye que los huevos de color oscuro, cáscara brillante y peso intermedio, puestos en meses lluviosos y almacenados por menos de 8 días presentan ventajas en la incubación. Asimismo, en sistemas donde no es necesario el apareamiento rotativo se recomienda usar el apareamiento continuo. Palabras clave: eclosión, incubación, Coturnix coturnix japonica, sistema de apareamiento, color de cáscara, brillo de cáscara. ABSTRACT With the aim to study the effect of some non genetic factors on fertility and hatchability of quails (Coturnix coturnix japonica) under two different reproduction systems (modalities of matching), three analyses of variance were realized assuming a binomial distribution of the data. There were used 2613 records for fertility (FE), 2178 for total hatchability (TH), and 1413 for fertile hatchability (FH) from a group of experimental quails. The effects of month of lay, generation, and type of matching were included for FE and for TH and FH were added egg weight, eggshell color and sheen. Water and food were offered ad libitum. The relation male: female was 1:3. Two modalities of reproduction were studied: rotary (rotating the male) and continuous (permanent presence of the male). Fertility, TH and FH averages were 78.7, 25.8, and 42.3%, respectively. The month of higher (P<0.01) fertility was March (91.1%), whereas November had the lowest (51.7%). The higher TH and FH (P<0.01) was in August (42.6 and 65.7%) and the lower TH and FH were observed in April (12.5 and 17.9%). There was higher fertility (P<0.01, 86.2%) in the eggs from continuous reproduction system. Eggs with higher TH (P<0.01) were those of brilliant eggshell (43.9%), dark color eggshell (48.0%) and intermediate weight (10.1 to 11.0 g; 44.0%). It can be concluded that the eggs of dark color, brilliant eggshell, and intermediate weight laid in the rainy months had better properties for the process of incubation. Likewise, in farms where the rotary matching is not necessary, it is advisable to use the continuous reproduction system. Keywords: hatchability, incubation, Coturnix coturnix japonica, reproduction system, eggshell color, eggshell sheen. INTRODUCCIÓN Pocos estudios se han realizado en Venezuela para determinar las potencialidades de las codornices y los factores que se deben considerar en el manejo integral de las granjas. En este sentido, se hace necesario recalcar la importancia que la fertilidad juega dentro de estos sistemas, particularmente en aquellas granjas de reproductoras, cuyo fin pasa por la producción de animales que irán posteriormente a la venta ó a producir huevos para el consumo humano. En estos sistemas es imprescindible obtener la mayor fertilidad posible, la cual a la postre redundará en un mayor porcentaje de eclosión (Kuurman et al., 2002). La expresión de la fertilidad, mortalidad y eclosión en las especies de aves domésticas tiene que ver con la carga genética de los animales y algunos factores ambientales que actúan independientes o en combinación para producir la respuesta reproductiva de estos (Kuurman et al., 2003). La forma y época de apareamiento, tiempo de almacenamiento de los huevos, peso del huevo, color y brillo de la cáscara son parámetros simples de tomar en cuenta para la mejora de la reproducción, pero en los sistemas de producción de codornices en Venezuela son poco estudiados y conocidos. El presente trabajo tiene como objetivo determinar la influencia del mes de postura, generación, tipo de apareamiento, peso del huevo, color y brillo de la cáscara sobre la fertilidad y eclosión de huevos de codornices. MATERIALES Y MÉTODOS Se utilizaron 2.613 registros de fertilidad, 2.178 para eclosión total y 1.403 para eclosión de huevos fértiles provenientes de experiencias realizadas en la Sección Laboratorio de Aves de la Facultad de Agronomía de la Universidad Central de Venezuela (UCV), la cual está ubicada en el Municipio Mario Briceño Iragorry del estado Aragua, a una altitud de 452 msnm. El área en estudio posee una temperatura promedio anual de 25,1ºC y una humedad relativa de 78,3%. Los huevos provenían de cuatro granjas de los estados Aragua y Yaracuy, los cuales se incubaron y se produjeron dos generaciones. Los animales resultantes de estas incubaciones se criaron y reprodujeron en la UCV, incubándose los huevos resultantes de los apareamientos de estos animales. Las variables evaluadas fueron huevos fértiles (%) y eclosión (%). Luego de ocurrir la eclosión se abrieron los huevos no eclosionados, de manera de apreciar la fertilidad de los mismos. Se consideraron huevos fértiles (además de aquellos que culminaron en la eclosión del polluelo) a los que presentaron embriones muertos (muerte en fase inicial o desarrollo temprano) y muertes en cascarón (muerte en fase final). De esta manera, la fertilidad se calculó como se describe a continuación:
Luego de la cría, los animales adultos (aproximadamente cuatro semanas de edad) se ubicaron en jaulas de alambre galvanizado con las siguientes dimensiones: 50 x 50 x 20 cm de largo, ancho y alto, respectivamente, para los animales en apareamiento continuo y 50 x 12,5 x 20 cm largo, ancho y alto, respectivamente, para los animales en apareamiento rotativo. Dichas jaulas fueron ubicadas en galpones con estructura de madera, cerrados con tela metálica en los laterales, techo de zinc y piso de cemento. A las aves en postura se ofreció un alimento comercial que contenía 15% de proteína y agua ad libitum. La relación macho:hembra en reproducción fue 1:3. El sistema de reproducción comprendió dos modalidades: rotativo, es decir, se colocaba al macho un día completo con cada hembra asignada, otorgándosele un día de descanso (cuarto día) entre cada ciclo reproductivo y continuo, en este caso el macho permanecía todo el tiempo en la misma jaula con las tres hembras. Durante el período de evaluación se registro la postura diaria (fechas) y el consumo de alimento (semanal). Diariamente los huevos fueron recogidos en la mañana y almacenados en un cuarto provisto con un equipo acondicionador de aire. La temperatura promedio en el cuarto de almacenamiento fue 21 ± 2ºC. Los huevos fueron incubados en un equipo marca Robinsón con capacidad para 3.900 huevos de gallina, a la cual se le realizó una adaptación en las bandejas para poder separar los huevos y obtener registros individuales de eclosión. En este sentido, las bandejas se subdividieron en pequeños recuadros con dimensiones de 5 x 6 cm. Al momento de la incubación se anotó el peso de cada huevo, el brillo y color de la cáscara, la fecha de incubación y el tiempo de almacenamiento. La temperatura promedio de incubación fue 36,5 – 37,5ºC y la humedad relativa entre 65 y 75%. Se realizaron incubaciones en los meses de marzo, abril, mayo, junio, agosto, noviembre y diciembre. Durante el período de postura, se aplicó un programa de iluminación artificial que consistió en ofrecer 5 h de luz adicionales, el cual se aplicó entre las 2 y las 7 am. Las evaluaciones se realizaron durante tres generaciones, a saber padres, primera y segunda generación. Se realizaron análisis de varianza asumiendo una distribución binomial en vista de que las variables estudiadas fueron registradas como 0 y 1 (Littell et al., 2002). Para detectar las diferencias entre medias se utilizó la prueba de t de Student. El modelo utilizado se describe a continuación: Yijkl = µ + Mpi + Genj + Apak + ℮ijkl donde: Yijkl : fertilidad de los huevos puestos en el mes i, provenientes de hembras de la generación j, sometidas a un tipo de apareamiento k, µ = media teórica de la población, Mpi = mes de postura (i = 3, 4, 5, 6, 8, 11 y 12), Genj = generación (j = 1, 2, 3), Apak = tipo de apareamiento (k = 1, 2) y ℮ijkl = residual con media (np) y varianza (npq), binomialmente distribuido. Los modelos para eclosión de los huevos fértiles y totales son similares al modelo descrito para fertilidad, adicionándose el efecto del peso del huevo. Los huevos se agruparon por peso en tres categorías: entre 5,0 y 10,0 g, entre 10,1 y 11,0 g y 11.1 g ó más. También se incluyeron los efectos de brillo y color de la cáscara. El brillo de la cáscara se clasificó en dos categorías, brillante y mate. Para el color de la cáscara se consideró la tonalidad de fondo y la presencia de manchas, clasificándose los mismos en tres categorías: claro (ausencia o muy poca pigmentación), medio (fondo claro con pocas manchas presentes) y oscuro (fondo más oscuro con abundantes manchas presentes, Figura 1). De igual manera, se incluyó el efecto del tiempo de almacenamiento; en este sentido, los huevos se agruparon de acuerdo a los siguientes criterios: entre cero y dos días de almacenamiento, entre tres y cinco días, entre seis y ocho días y entre nueve y quince días de almacenamiento. Asimismo los efectos de generación y tipo de apareamiento se excluyeron de los análisis para eclosión. Los promedios y sus errores estándar asintóticos para ambos caracteres se calcularon utilizando las fórmulas siguientes (Littell et al., 2002):
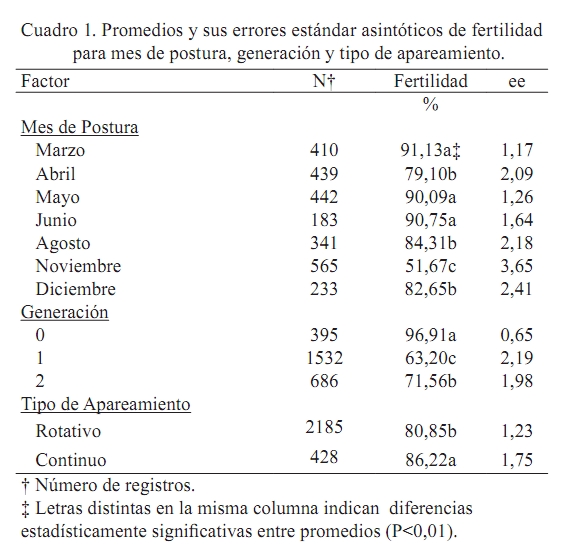
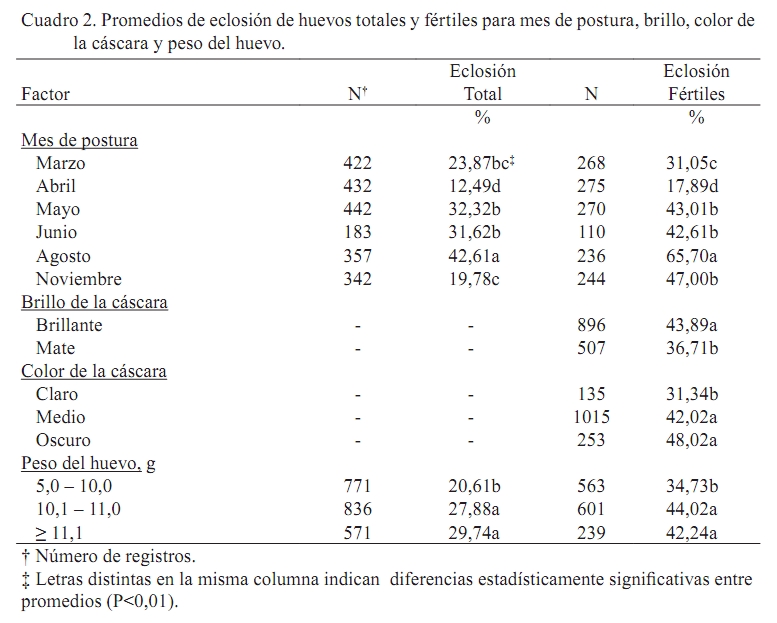
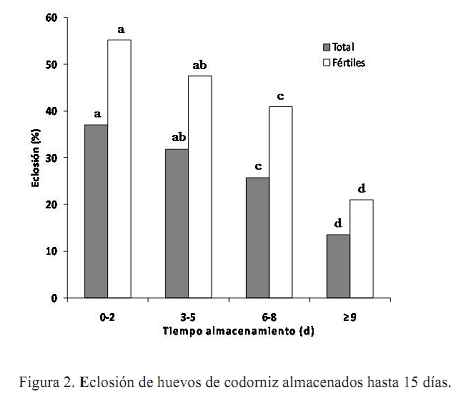
Error estándar asintótico = promedio x (1 – promedio) x (error estándar del estimado). RESULTADOS Y DISCUSIÓN Fertilidad El promedio ajustado correspondió a 78,72% con un error estándar de 1,32%. Los promedios encontrados en el presente estudio fueron superiores en 17% a los señalados en la literatura por Martínez (1990). Sin embargo, estos promedios están entre 2 y 12 puntos por debajo de los promedios reportados por Lucotte (1990), Espidea (1995), Morantes (1995), Quintana (1999), Lembcke et al. (2001), Mejia (2005) y Ravel (2006). Es evidente que pautas de manejo y las diferencias en el material genético entre una y otra investigación pudieran estar introduciendo variación en los resultados. Esta aseveración fue hecha con anterioridad por Martínez (1990), quien señaló que la baja calidad genética de las poblaciones de codornices presentes en la zona central de Venezuela pueda ser la causa de los bajos niveles de fertilidad observados. La baja calidad genética de los animales probablemente sea consecuencia de la ausencia de granjas especializadas en reproducción en el país, sumado al hecho de que cada granja productora de huevos incuba algunos de estos para obtener reposición y para la venta, sin seguir estrictos programas de reproducción y selección. Los análisis reflejaron efecto significativo (P<0,01) del mes de postura, generación y tipo de apareamiento. Los meses de mayor fertilidad corresponden a marzo, mayo y junio, en los cuales el porcentaje de fecundidad fue superior al 90% (Cuadro 1). Abril, agosto y diciembre fueron meses intermedios respecto al carácter estudiado, con valores que oscilaron entre 79 y 84%, mientras que el peor mes fue noviembre, donde el porcentaje de huevos fértiles se ubicó en 51%. Se evidencia una disminución de la fertilidad en el mes de mayor temperatura ambiental y menor humedad relativa (Abril, Cuadro 1), lo cual coincide con el reporte de Ozbey y Ozcelik (2004). Probablemente la causa de la menor fertilidad tiene que ver con una reducción del consumo de alimento por las causas ambientales anteriormente mencionadas, que redunda negativamente sobre la reproducción. Asimismo, se nota una disminución drástica de la fecundidad en el mes de noviembre, probablemente motivada por aumentos excesivos de la humedad relativa durante estos meses, situación que se expresa en una menor actividad sexual de los machos y, por ende, desencadena en reducción de la fertilidad de los huevos incubados. Esta relación entre condiciones ambientales (temperatura y humedad relativa) fue señalada por Lucotte (1990) y Vargas (2005) como un punto crucial a considerar en los sistemas de producción con codornices. El mayor porcentaje de fertilidad (P<0,01) se obtuvo en la generación inicial (generación 0, Cuadro 1) con un valor de 97%, seguido por la generación dos con un valor de 72% y el menor porcentaje de fertilidad corresponde a la generación uno, con un valor de 63%. Es necesario mencionar que en esta población la selección sólo se ha basado en aspectos morfológicos: ausencia de defectos anatómicos, enfermedades detrimentales hereditarias, entre otros. A la par se continúa la fase evaluativa que comprende la determinación de los parámetros genéticos para los caracteres de interés económico, que a la postre, servirán como indicativo para diseñar los planes de mejoramiento genético (selección y cruzamiento). Vale la pena decir que es muy probable que la generación enmascare algunos problemas de manejo que pudieran estar reduciendo la fertilidad tanto en machos como en hembras, siendo este el caso de la primera generación. Empero, esa reducción es recuperada con el aumento de la fecundidad observado en la segunda generación. Ravel (2006) observó un comportamiento similar. En este caso, existe una disminución clara de la fertilidad a medida que pasan las generaciones, atribuyéndose este efecto, según el autor, a un aumento de la edad de los reproductores enmascarado dentro de la generación. Se obtuvo la mayor fertilidad en los animales sometidos a apareamientos continuos (Cuadro 1). Para este caso particular, en promedio, los huevos provenientes de hembras que están continuamente con el macho fueron 6% (P<0,01) más fértiles que sus homólogos provenientes de hembras sometidas al sistema de apareamiento rotacional. Este resultado es similar al reporte de Mejia (2005) quién observó diferencias de hasta 7% de fertilidad a favor de los huevos provenientes de apareamientos continuos. Es probable que el apareamiento continuo del macho con las distintas hembras asegure la mayor fertilidad de los huevos provenientes de este sistema. Caso contrario en el sistema rotatorio, en el cual muchos de los apareamientos quizás no sean efectivos debido al estrés causado a los animales al retirar el macho de la jaula y colocarlo en una jaula con otra hembra. En todo caso, la utilización de uno u otro sistema de apareamiento dependerá de los objetivos de la unidad productiva. Para los fines de esta investigación, aún cuando el sistema de apareamiento rotacional resultó en menor fertilidad, se justifica este manejo reproductivo, pues el objetivo final es el mejoramiento genético de la población, objetivo que necesariamente requiere de la identificación individual y precisa de los machos y hembras en reproducción y, por supuesto, de sus hijos (as). Eclosión Se obtuvieron promedios de eclosión total y de huevos fértiles de 25,8 y 42,3%, con errores estándar de 2,0 y 2,2%, respectivamente. El promedio de eclosión para huevos fértiles fue inferior a los reportes de Ortega (1994) y Espidea (1995), pero superior al señalado por Mejía (2005), Uztariz (2005) y Ravel (2006). Es necesario mencionar que dichas diferencias pueden tener causas diversas, las cuales se refieren no sólo a la temperatura, humedad y movimiento de los huevos, sino que se debe adicionar el tiempo de almacenamiento de los huevos, las condiciones de almacenamiento y el peso del huevo. Estas condiciones pueden variar entre una investigación y otra, siendo fuentes silentes y no identificadas de variación (Hassam et al., 2005). El Cuadro 2 muestra los valores de eclosión de los huevos totales y fértiles, de acuerdo al mes de postura. En el mismo se observa la superioridad en los valores de eclosión en ambos casos para el mes de agosto, el cual correspondió a 42,6 y 65,7%, respectivamente (P<0,01). Asimismo, en ambos casos el mes que presentó la menor eclosión fue abril (P<0,01), con valores de 12,5 y 17,9% para la eclosión total y de los huevos fértiles, respectivamente. Resultados disímiles son reportados por Baratta (1993). Al igual que para el caso de fertilidad, tanto abril como noviembre presentaron los menores valores de eclosión total (Cuadro 1 y Cuadro 2), observándose una correspondencia de los meses en ambas características. Tal y como se señaló con anterioridad, la dificultad para controlar las condiciones internas durante la incubación puede estar influenciando los resultados obtenidos en meses críticos. De esta manera se expresa la reducción en el mes de abril, cuando las temperaturas son elevadas durante el día y ocurren reducciones drásticas en ésta variable ambiental en horas nocturnas. Situación similar fue observada en el mes de noviembre, cuando la humedad relativa, tanto ambiental como dentro de la incubadora fue elevada, ocasionando un funcionamiento inadecuado de la misma, lo que condujo a una disminución de la eclosión. Por otra parte, es menester mencionar que tanto en abril como en noviembre se obtuvieron los menores valores de fertilidad, lo que pudiera estar influenciando el resultado de eclosión total, puesto que es probable que se estén incubando muchos huevos, los cuales no son fértiles. Los huevos de cáscara brillante eclosionaron 7% más (P<0,05) que los huevos de cáscara mate (Cuadro 2). Resultados similares han sido reportados por Quintana (1999). Es posible que los huevos brillantes mantengan un menor intercambio gaseoso y de humedad con el medio ambiente que los huevos mate, debido a la presencia de la película protectora que los recubre. En consecuencia, en estos huevos se conserva una mejor integridad del albumen, que ha sido señalado como un mejor equilibrio ácido – base de este, situación que resulta en la sobrevivencia del embrión, y por ende, en la eclosión (Reis et al., 1997; Lapao et al., 1999). Dentro de la categoría del color de la cáscara del huevo, los resultados reflejan ventaja en eclosión (P<0,01) de los huevos cuya cáscara tiene mayor cantidad de manchas y estás son de mayor tamaño (huevos oscuros, Cuadro 2). Estos huevos mostraron 48% de eclosión, mientras que en segundo lugar se ubicaron los que poseen manchas intermedias sobre fondo claro (medio) con 42% de eclosión y los que menos eclosionaron (31%) fueron aquellos cuya cáscara poseía escasas manchas (claros). Investigaciones realizadas con anterioridad (Lucotte, 1990; Quintana, 1999) señalan la superioridad en eclosión de los huevos con mayor número de manchas, atribuyéndose este efecto a que los huevos con mayor número de manchas permanecen por más tiempo en el tracto reproductivo de la hembra, aumentando la protección de este (mayor grosor y cantidad de cutina sobre los poros) contra los agentes patógenos que pudieran interferir con el desarrollo embrionario. Para el caso particular del peso, los huevos que más eclosionaron (P<0,01) fueron aquellos cuyo peso osciló entre 10,1 y 11,0 g (44%) y en segundo lugar los huevos que pesaron más de 11,1 g, siendo los huevos más livianos (5,0 - 10 g) los que menos eclosionaron (Cuadro 2). Estos resultados coinciden con el reporte de Lucotte (1990) quién evidenció mayores porcentajes de eclosión en los huevos de peso intermedio (alrededor de 10 g). Una explicación probable se basa en el hecho de que los huevos de peso intermedio poseen una alta capacidad de intercambio de agua y gases con el medio ambiente, debido a la presencia de un mayor número de poros en la cáscara (González et al., 1999). Este intercambio con el medio ambiente ha sido relacionado directamente con una mayor calidad del albumen, lo cual asegura la sobrevivencia del embrión (Reis et al., 1997; Lapao et al., 1999). Asimismo, se ha señalado que los huevos grandes tienen menos poros, lo que dificulta el intercambio de gases y agua con el medio ambiente que rodea al huevo; con la consecuente reducción de la pérdida de peso del huevo y reducción en la calidad del albumen que conduce a la muerte del embrión (Hassan et al., 2005). La Figura 2 muestra los valores de eclosión de huevos totales y fértiles de acuerdo al tiempo de almacenamiento de los mismos antes de la eclosión. Claramente se observa un descenso de los porcentajes de eclosión en ambos casos a medida que aumenta el tiempo de almacenamiento. La mejor respuesta (P<0,01) se obtuvo con un tiempo de almacenamiento de hasta dos días (37,0 y 55,2% para eclosión de huevos totales y fértiles, respectivamente), para luego producirse un descenso gradual en los porcentajes de eclosión hasta el tercer período (6 – 8 días), luego del cual ocurre una disminución drástica de la eclosión (hasta 20%) en el último período que incluye huevos almacenados entre 9 y 15 días. Los resultados coinciden con los reportes de González et al. (1999), Fasenko et al. (2001a,b), Kuurman et al. (2002), Hassan et al. (2005) y Uztariz (2005). Quizás la causa del comportamiento observado tenga que ver con el deterioro de las condiciones internas del huevo, principalmente pérdida de humedad y aumento del pH al aumentar los días de almacenamiento, lo cual conduce a la licuefacción del albumen, perdiendo este su capacidad antimicrobial. También se ha señalado que el albumen posee una función importante para mantener la posición correcta del huevo, interactuando con las membranas internas del huevo, estableciéndose la teoría de que el tiempo de almacenamiento del huevo reduce las posibilidades de una eclosión efectiva, al aumentar las posibilidades de muerte del embrión por las causas antes descritas (Brake et al., 1997). CONCLUSIONES No se observó una tendencia marcada de fertilidad respecto a los meses estudiados; sin embargo, se evidencia una época superior para incubación, debido a que los huevos puestos entre mayo y agosto para ser incubados presentaron el mayor porcentaje de fertilidad y eclosión. Asimismo, cuando el sistema no requiera la identificación individual de los reproductores y sus hijos es aconsejable implementar el apareamiento continuo, puesto que la fertilidad es superior. Por otra parte, los huevos a ser incubados deben ser de tamaño intermedio (10,0 – 11,0 g), de cáscara brillante, de coloración oscura y no deben ser almacenados para su posterior incubación por más de ocho días, para de esta manera asegurar un mayor porcentaje de eclosión. LITERATURA CITADA
Copyright © 2009 - Zootecnia Tropical The following images related to this document are available:Photo images[zt09002f1.jpg] [zt09002f2.jpg] [zt09002c2.jpg] [zt09002c1.jpg] |
| |||||||||


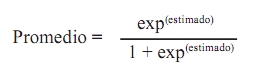
{kind=link}
{kind=link}
{kind=link}
{kind=link}