 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
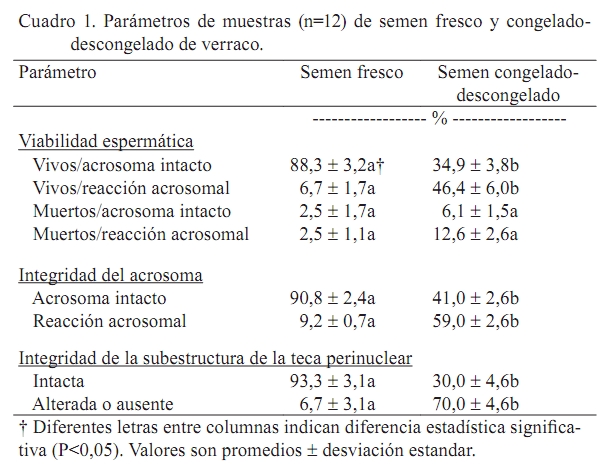
Zootecnia Tropical, Vol. 27, No. 1, 2009, pp. 17-24 Artículos Científicos Alteraciones en la integridad del acrosoma y de la teca perinuclear en semen criopreservado de verraco Alterations in the integrity of the acrosome and perinuclear theca in cryopreserved boar spermatozoa Manuel Barrientos Morales1,2, María De L. Juárez Mosqueda2, María E. Trujillo Ortega3 y Felipe Montiel Palacios1* 1 Departamento de Reproducción, Facultad de Medicina Veterinaria y Zootecnia, Universidad Veracruzana, Circunvalación S/N Esq. Yañez, C.P. 91710, Veracruz, Veracruz. México. *Correo electrónico: fmontiel@uv.mx Recibido: 1/05/2008 Aceptado: 24/10/2008 Code number: zt09003 RESUMEN El objetivo del estudio fue investigar el efecto de la criopreservación sobre la integridad del acrosoma y de la subestructura de la teca perinuclear (PTS) del espermatozoide de verraco. Se utilizó semen proveniente de seis verracos. El semen se colectó, se diluyó y se determinó la motilidad y concentración espermática, incluyendo en el estudio sólo los eyaculados con motilidad ≥70%. El semen diluido se dividió en dos muestras: una para evaluarse en fresco y otra para evaluarse después de la congelación-descongelación. Ambos tipos de muestras se utilizaron para evaluar la viabilidad espermática y la integridad del acrosoma mediante microscopía de luz usando la técnica de triple tinción y para evaluar la integridad de la PTS mediante microscopía electrónica. El semen congelado-descongelado tuvo mayor proporción (P<0,05) de espermatozoides vivos con reacción acrosomal, comparado con el semen fresco. El proceso de congelamiento-descongelamiento redujo (P<0,05) la proporción de espermatozoides con PTS intacta. El valor de espermatozoides con reacción acrosomal se correlacionó con la proporción de espermatozoides con PTS alterada o ausente, tanto en semen fresco (r=0,93; P<0,05) como en semen congelado-descongelado (r=0,71; P<0,05). En conclusión, el semen criopreservado de verraco presentó alteraciones en la integridad del acrosoma y de la PTS que pudieron ser debidas al fenómeno de criocapacitación. Palabras clave: criopreservación, espermatozoide de verraco, reacción acrosomal, teca perinuclear. ABSTRACT The objective of this study was to investigate the effect of cryopreservation on the integrity of the acrosome and the perinuclear theca substructure (PTS) of boar spermatozoa. Semen from six boars was used. The semen was collected, diluted, and sperm motility and concentration determined. Only ejaculates showing ≥70% motility were used in this study. Diluted semen was divided in two samples: one to be evaluated fresh, and other to be evaluated after freezing-thawing. Fresh and frozen-thawed semen samples were used to assess sperm viability and acrosome integrity by light microscopy using the triple staining technique and to evaluate the PTS integrity by electron microscopy. The frozen-thawed semen had a greater proportion (P<0.05) of acrosome reacted live spermatozoa, compared to the fresh semen. The freezing-thawing process reduced (P<0.05) the proportion of intact PTS spermatozoa. The value of acrosome reacted spermatozoa was correlated with the proportion of spermatozoa with altered or absent PTS, both in fresh (r=0.93; P<0.05) and in frozen-thawed (r=0.71; P<0.05) semen. In conclusion, cryopreserved boar semen showed some alterations in the integrity of the acrosome and the PTS that might be due to cryocapacitation. Keywords: acrosome reaction, boar spermatozoa, cryopreservation, perinuclear theca INTRODUCCIÓN La criopreservación daña la membrana plasmática y la membrana acrosomal externa del espermatozoide (De Leeuw et al., 1990; McGann et al., 1998). Después del descongelamiento, una pequeña población de los espermatozoides que sobreviven a este procedimiento muestra signos de criocapacitación que se refiere a cambios en los patrones de movilidad, redistribución del Ca2+, etc., que asemejan el proceso de capacitación (Watson, 1995; Bailey et al., 2000). La criocapacitación resulta en la ocurrencia de una versión similar pero desorganizada de la reacción acrosomal (Watson, 2000). En el espermatozoide porcino criopreservado, el proceso de enfriamiento y posterior re-calentamiento induce cambios similares a la capacitación (Kaneto et al., 2002), que son parcialmente responsables de la baja fertilidad general del semen congelado de verraco (Watson, 2000; Green y Watson, 2001). Por otro lado, el citoesqueleto participa en el soporte de la membrana plasmática y de la membrana acrosomal y algunos de sus elementos son termosensibles (Omega et al., 2006). El principal elemento del citoesqueleto de la cabeza del espermatozoide es la teca perinuclear (PT) (Longo et al., 1987). La PT es una cápsula rígida que cubre el núcleo del espermatozoide de mamíferos y tiene como función la unión de membranas espermáticas (Korley et al., 1997) y la preservación de su integridad (Watson, 2000). La PT está subdividida en dos regiones: las capas subacrosomal y postacrosomal (Omega et al., 2006). La capa postacrosomal se cree que participa en la activación del ovocito durante la fertilización (Sutovsky et al., 1997) y es el sitio para la actina en los espermatozoides de algunos mamíferos (Yagi y Paranko, 1995; Mujica et al., 2003). En la región apical de la capa postacrosomal del espermatozoide del toro, carnero y conejillo de Indias se ha detectado una subestructura, llamada subestructura de la teca perinuclear (PTS) (Olson et al., 1983; Juárez Mosqueda y Mujica, 1999). En el espermatozoide del conejillo de Indias, la F-actina está involucrada en la estabilización de la PTS (Juárez Mosqueda y Mujica, 1999). En el espermatozoide del verraco, la actina participa en la regulación del volumen (Petrunkina et al., 2004). La localización de la actina en la región acrosomal de algunas especies de mamíferos (Camatini et al., 1986; Flaherty et al., 1988; Yagi y Paranko, 1995) respalda su posible participación en la capacitación espermática y en la reacción acrosomal (RA); así, la polimerización y despolimerización de la actina pueden estar involucradas en la función espermática (Breitbart et al., 2005). En el espermatozoide del cerdo, conejillo de Indias, toro, ratón y carnero, la polimerización de la actina ocurre durante la capacitación (Castellani Ceresa et al., 1993; Brener et al., 2003; Cabello Agüeros et al., 2003) y el desdoblamiento de la F-actina debe ocurrir para que se lleve a cabo la RA (Brener et al., 2003). En el espermatozoide del verraco, la inhibición de la polimerización de la actina bloquea su capacidad para fertilizar in vitro (Castellani Ceresa et al., 1993). En otros tipos de células, la despolimerización prematura de la F-actina es causada por el enfriamiento (Hall et al., 1993; Saunders y Parks, 1999). El espermatozoide del verraco se ve dañado severamente durante el congelamiento (Maldjian et al., 2005) y no más del 50% sobrevive a la criopreservación (Eriksson y Rodríguez Martínez, 2000; Carvajal et al., 2004). El daño a la PT durante el proceso de congelamiento-descongelamiento ha sido reportado en el espermatozoide del toro (Omega et al., 2006), pero sólo pocos estudios han documentado el daño que ocasiona la criopreservación en la PT en el verraco. El objetivo del presente estudio fue investigar el efecto de la criopreservación sobre la integridad del acrosoma y de la PTS del espermatozoide de verraco. MATERIALES Y MÉTODOS Colección de semen y evaluación Se utilizaron seis verracos maduros de raza Duroc de probada fertilidad, mantenidos bajo las mismas condiciones de manejo y alimentación. Usando el método de la mano enguantada, de cada verraco se recolectaron dos eyaculados con intervalo de una semana, obteniendo en total 12 eyaculados para el estudio. De cada eyaculado se colectó la fracción rica en espermatozoides y cada eyaculado fue procesado de manera separada. El semen se diluyó (1:1 [v:v]) con diluyente MR-A® (Kubus®, Madrid, España) y se determinaron la motilidad y concentración espermáticas mediante técnicas de laboratorio de rutina. Todos los eyaculados usados en el estudio tuvieron ≥70% de motilidad. Para llevar a cabo la evaluación de la integridad del acrosoma y de la PTS el semen diluido se dividió en dos muestras: una para ser evaluada en fresco y la otra para ser evaluada después del proceso de congelamiento-descongelamiento. Protocolo de congelamiento y descongelamiento del semen Para el congelamiento del semen, las muestras fueron procesadas de acuerdo al protocolo de Bwanga (1990). Brevemente, el semen diluido fue enfriado (Coolmake®, Minitube®, Cd. de México, México) a 15ºC por 2 h y después centrifugado (1.200 rpm/3 min). Para remover el diluyente, los pellets con espermatozoides se lavaron tres veces en solución salina (0,9% NaCl [p/v], osmolaridad de 308 mOsm/L) y fueron centrifugados (1.200 rpm/3 min). Después, los pellets con espermatozoides fueron re-diluidos con un segundo diluyente (DII, 80 mL de lactosa al 11% + 20 mL de yema de huevo, pre-enfriados a 15ºC) para producir una concentración espermática de 1,5 x 109/mL. Los espermatozoides fueron enfriados a 5ºC por 90 min y mezclados con un tercer diluyente (DIII, 89,5 mL de DII + 1,5 mL de Equex STM [Nova Chemicals Sales Inc., Scituate, MA, USA] + 9 mL de glicerol, pre-enfriados a 5ºC) para producir una concentración final para ser congelada de 1 x 109 espermatozoides/mL y 3% de glicerol. El semen fue empacado en pajillas francesas de 0,5 mL (Minitube®, Cd. de México, México), que fueron congeladas mediante exposición a vapores de nitrógeno líquido (NL; -130 a –150°C) por 20 min y después sumergidas en NL a –196ºC para su almacenamiento. El descongelamiento de las pajillas se hizo en baño María a 37°C por 20 seg. Inmediatamente después del descongelamiento, los espermatozoides fueron usados para evaluar la integridad del acrosoma y de la PTS. Evaluación de la viabilidad de los espermatozoides y de la integridad del acrosoma Se usó semen fresco y semen congelado-descongelado para evaluar la proporción de espermatozoides vivos, muertos y con reacción acrosomal en cada muestra, según la técnica de tinción triple descrita por Talbot y Chacón (1981). Brevemente, una alícuota de 100 µL de semen se mezcló con 100 µL de azul tripán al 2%, se incubó a 37ºC por 15 min, se centrifugó (1.200 rpm/3 min) y el pellet con espermatozoides se resuspendió en 2 mL de agua destilada y se centrifugó dos veces. Después, el pellet con espermatozoides se fijó en 1 mL de glutaraldehído al 3% a 4ºC por 30 min, se lavó en agua destilada, se centrifugó (1.200 rpm/3 min) y se resuspendió en solución salina fosfato-buferada Dulbecco (PBS, compuesta de 136,89 mM de NaCl, 2,68 mM de KCl, 8,1 mM de Na2HPO4 y 1,46 mM de CaCl2-2H2O) al volumen inicial. Entonces, una gota de 10 a 20 µL de suspensión de espermatozoides se esparció en un portaobjetos de cristal. El portaobjetos se sumergió en solución de café Bismark al 8% en agua destilada a 37ºC por 15 min, se lavó con agua, se secó al aire, se tiñó con solución de rosa de Bengala al 0,8% por 1 min a temperatura ambiente, se lavó con agua y se examinó con un microscopio de luz (X 1000). Se examinaron un total de 100 espermatozoides por portaobjetos para evaluar la viabilidad espermática y la integridad del acrosoma. Se observaron los siguientes patrones de tinción: 1) rosa en la región acrosomal y café en la región post-acrosomal en espermatozoides vivos con acrosoma intacto (EVAI), 2) sin tinción en la región acrosomal y café en la región post-acrosomal en espermatozoides vivos con reacción acrosomal (EVRA), 3) rosa en la región acrosomal y azul en la región post-acrosomal en espermatozoides muertos con acrosoma intacto (EMAI) y 4) sin tinción en la región acrosomal y azul en la región post-acrosomal en espermatozoides muertos con reacción acrosomal (EMRA). Evaluación de la integridad de la PTS Se utilizó semen fresco y semen congelado-descongelado para la evaluación de la morfología de la PT mediante microscopía electrónica (Ursitti y Wade, 1993). Los espermatozoides se lavaron en solución salina y se ajustaron a una concentración de 35 x 106 células/mL. Para exponer la superficie de la PT, la membrana plasmática, la membrana nuclear y el acrosoma se solubilizaron agregando el detergente no iónico Brij 36-T (Canamex, Nuevo León, México) a una concentración final de 1,2% y los espermatozoides se incubaron por 5 min a temperatura ambiente. Las muestras se centrifugaron (2.500 rpm/3 min), se lavaron en agua destilada y se fijaron en fijador Karnowsky por 20 min a temperatura ambiente. Después, las muestras fueron centrifugadas (2.500 rpm/3 min), lavadas dos veces en PBS y dos veces en agua destilada y resuspendidas en agua destilada al volumen inicial. Una gota de cada suspensión de espermatozoides se colocó en una rejilla cubierta con colodión y carbón, dejándola 5 a 10 min para permitir a los espermatozoides adherirse a la rejilla. Las muestras se tiñeron con solución acuosa de 0,02% de fosfotungstato (Merck, Darmstadt, Germany) por 3 min, y fueron enjuagadas poniendo una gota de agua destilada en la rejilla. Las rejillas se colocaron sobre papel filtro en cajas de Petri y se cubrieron para secar. Las muestras fueron examinadas en un microscopio de transmisión electrónica Zeis EM-9 (Oberkochen, Germany) a 50 kv. Para evaluar la integridad de la PTS, 100 espermatozoides se evaluaron y clasificaron y la característica considerada para evaluar la condición de la PT fue la apariencia morfológica de la subestructura sobre la capa post-acrosomal. Las muestras fueron clasificadas como: 1) intactas o normales, cuando la PTS apareció como una fila continua de papilas, y 2) alterada o ausente, cuando la PTS tuvo algún daño, o las células no presentaron esta estructura, respectivamente (Omega et al., 2006). Análisis estadístico Las diferencias entre semen fresco y congelado-descongelado para la proporción de EVAI, EVRA, EMAI, EMRA, espermatozoides con PTS intacta y espermatozoides con PTS alterada o ausente se analizaron mediante análisis de varianza, considerando un nivel de significancia estadística de P<0,05. Se utilizó la correlación de Pearson para analizar la correlación entre la integridad del acrosoma y la integridad de la PTS. Se utilizó el paquete estadístico SAS versión 6.08 para Windows (SAS, 1988). RESULTADOS El semen congelado-descongelado tuvo menor proporción (P<0,05) de EVAI y mayor proporción (P<0,05) de EVRA, comparado con el semen fresco (Cuadro 1). No hubo diferencia (P>0,05) en el porcentaje de EMAI o EMRA en el semen fresco y congelado-descongelado (Cuadro 1). La proporción general de espermatozoides con acrosoma intacto (vivos y muertos) fue mayor (P<0,05) en el semen fresco (Cuadro 1). El proceso de congelamiento-descongelamiento redujo (P<0,05) la cantidad de espermatozoides con PTS intacta, comparados con los espermatozoides evaluados en fresco (Cuadro 1). Se encontró relación entre la integridad del acrosoma y la integridad de la PTS. La proporción de espermatozoides con reacción acrosomal se correlacionó con la proporción de espermatozoides con PTS alterada o ausente, tanto en el semen fresco (r=0,93), como en el congelado-descongelado (r=0,85). En el semen fresco, la proporción de espermatozoides con PTS intacta se correlacionó con la proporción de EVAI (r=0,88) y con la proporción de espermatozoides con acrosoma intacto (r=0,91), mientras que la proporción de espermatozoides con PTS alterada o ausente tuvo muy alta correlación con la proporción de EVRA (r=0,95) y se correlacionó con los espermatozoides con reacción acrosomal (r=0,91). En el semen congelado-descongelado, la proporción de espermatozoides con PTS intacta se correlacionó con la proporción de EVAI (r=0,86) y tendió a correlacionarse con la proporción de espermatozoides con acrosoma intacto (r=0,69), mientras que la proporción de espermatozoides con PTS alterada o ausente tendió a correlacionarse con las proporciones de espermatozoides con reacción acrosomal (r=0,69) y EVRA (r=0,61). DISCUSIÓN El espermatozoide del verraco es particularmente susceptible al choque por frío durante la criopreservación, resultando en extensivo daño a la membrana y consecuentemente, disminución en la fertilidad del semen congelado (Chatterjee y Gagnon, 2001; Maldjian et al., 2005). Dado que el acrosoma contiene las enzimas necesarias para el espermatozoide para penetrar y fertilizar el ovocito (Eddy y O’Brien, 1994), a menor daño acrosomal durante la criopreservación, mayor tasa de fertilidad resultante (Zeng y Terada, 2001). En el presente estudio, el mayor porcentaje de EVAI en el semen fresco, comparado con el semen congelado-descongelado, mostró que la criopreservación afectó la integridad del acrosoma; esto podría explicar en parte las bajas tasas de fertilidad que se obtienen al utilizar semen criopreservado de verraco. La capacitación espermática y la RA son dos pasos clave en el proceso de fertilización (Gadea et al., 2005). La capacitación del espermatozoide puede ser inducida in vitro en presencia de diferentes medios (Yanagimachi, 1994). La RA ocurre cuando el espermatozoide ha sufrido la capacitación (Spungin et al., 1995). Sin embargo, en el espermatozoide del verraco el enfriamiento lento a 5ºC antes del congelamiento indujo cambios en la membrana, similares a los que se presentan durante la capacitación (Maxwell y Johnson, 1997; Green y Watson, 2001). Se ha sugerido que los espermatozoides sufren un proceso continuo de desestabilización de la membrana durante el congelamiento y el descongelamiento, llevando finalmente a muerte celular, lo que podría explicar la alta proporción de espermatozoides con RA presente en el semen descongelado de verraco (Cremades et al., 2005). Esto fue comparable al resultado de nuestro estudio, ya que el porcentaje de espermatozoides con RA fue mayor en el semen congelado-descongelado que en el semen fresco. La despolimerización prematura de la F-actina como resultado del enfriamiento (Hall et al., 1993; Saunders y Parks, 1999) podría contribuir a una fusión desorganizada de las membranas espermáticas después del congelamiento (Watson, 2000). Omega et al. (2006) reportaron que la criopreservación dañó la PT del espermatozoide del toro, similar a nuestros hallazgos en el espermatozoide del verraco. Esto apoya la propuesta de diversos investigadores de que la disminución en la viabilidad del semen criopreservado puede atribuirse no sólo a daño primario en la membrana plasmática (McGann et al., 1988; Jager et al., 1990; Holt, 2000), sino también al daño a elementos internos, tales como el citoesqueleto (Watson, 2000), como fue observado por Omega et al. (2006) en el espermatozoide del toro. Sin embargo, no hay reportes de estudio similares realizados en cerdos. En el presente estudio, la criocapacitación pudo haber alterado la integridad de la PTS, ya que la proporción de espermatozoides con PTS alterada o ausente fue mayor en los espermatozoides congelados-descongelados, comparados con los espermatozoides frescos. Omega et al. (2006) demostraron que la PTS del espermatozoide del toro podía usarse como un marcador morfológico para evaluar la integridad de la PT. Esta misma hipótesis se planteo en el presente estudio con espermatozoides de verraco. Similar al reporte en toros (Omega et al., 2006), en cada muestra de semen de nuestro estudio se pudieron detectar alteraciones en la PTS, sin importar si se trataba de espermatozoides frescos o congelados-descongelados y se encontró una mayor proporción de espermatozoides con PTS alterada o ausente en los espermatozoides congelados-descongelados. En el espermatozoide del conejillo de Indias, el trastorno en la F-actina lleva a pérdida de la PTS (Juárez Mosqueda y Mújica, 1999). En el espermatozoide del toro, el congelamiento-descongelamiento puede resultar en cambios en el citoesqueleto de microfilamentos que compone la zona base de la PTS, resultando en pérdida parcial o completa de la PTS (Omega et al., 2006). En el espermatozoide criopreservado de toro, la integridad del acrosoma, más que la motilidad espermática, tuvo un efecto significativo en las tasas de fertilización in vitro (Tartaglione y Ritta, 2004). Algunos autores han reportado la participación de la PT en el mantenimiento y formación de los dominios de la membrana plasmática funcional de la cabeza del espermatozoide, la estabilidad del núcleo y la activación del ovocito (Longo y Cook, 1991). El daño a la integridad de la PT podría afectar otras funciones fisiológicas del espermatozoide y tiene también un papel importante en la baja viabilidad del semen congelado-descongelado (Omega et al., 2006). Existen pocos reportes de estudios realizados sobre alteraciones en la PT del espermatozoide del cerdo debidas a la criopreservación. Nuestro estudio demostró que la criopreservación dañó la integridad del acrosoma y de la PTS del espermatozoide del verraco, lo que podría ser una de las causas de la baja fertilidad obtenida al utilizar semen congelado de verraco. CONCLUSIONES La criopreservación del espermatozoide del verraco afectó la integridad del acrosoma y de la PTS, probablemente debido a la criocapacitación. Sin embargo, se deben realizar estudios para saber si los espermatozoides congelados-descongelados que sufren RA debido a la criocapacitación son capaces de fertilizar al óvulo. Las alteraciones en la integridad del acrosoma y de la PTS podrían ser responsables de las bajas tasas de fertilidad obtenidas al utilizar semen congelado-descongelado de verraco. AGRADECIMIENTOS El presente proyecto de investigación fue financiado por el programa CGAP-UNAM (PAPIIT IN-206506). LITERATURA CITADA
Copyright © 2009 - Zootecnia Tropical The following images related to this document are available:Photo images[zt09003c1.jpg] |
| |||||||||

{kind=link}