 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
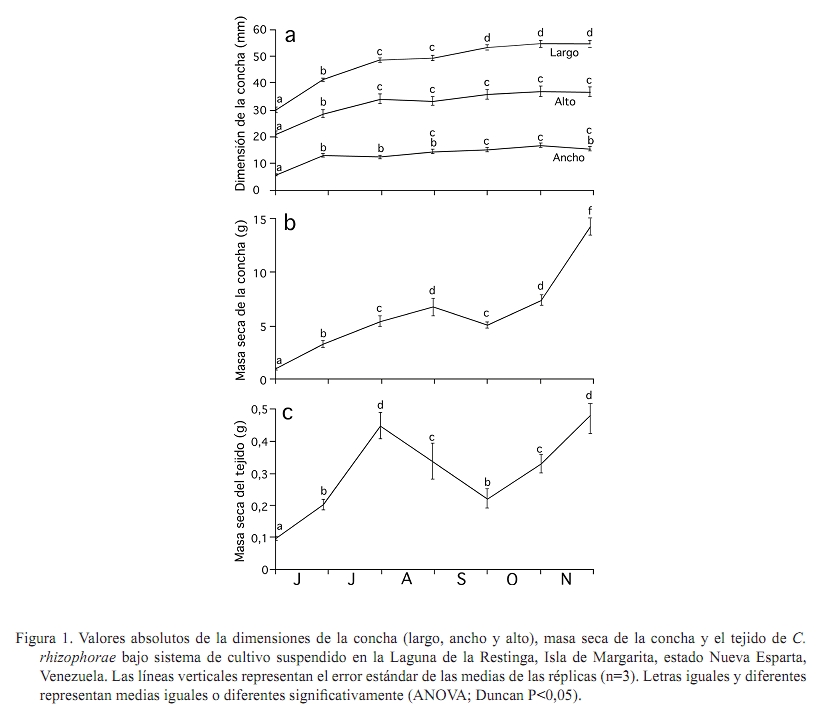
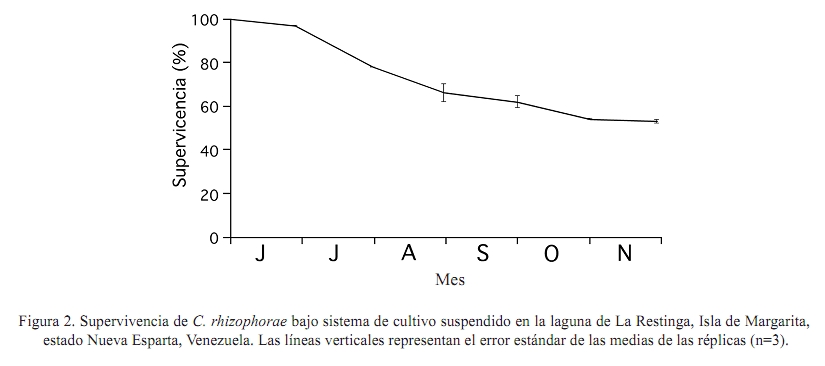
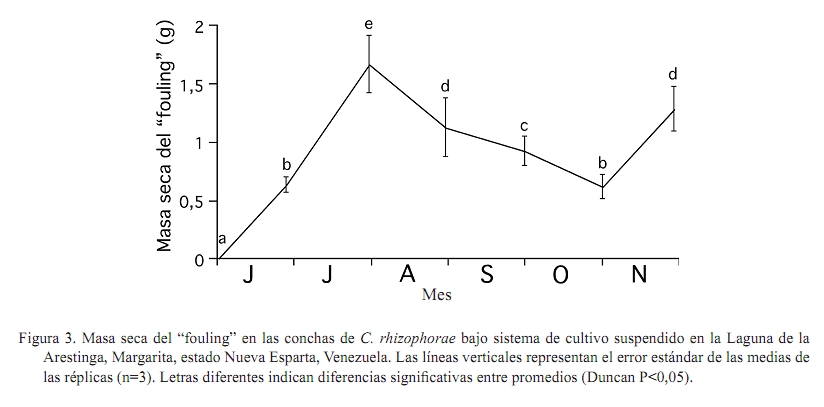
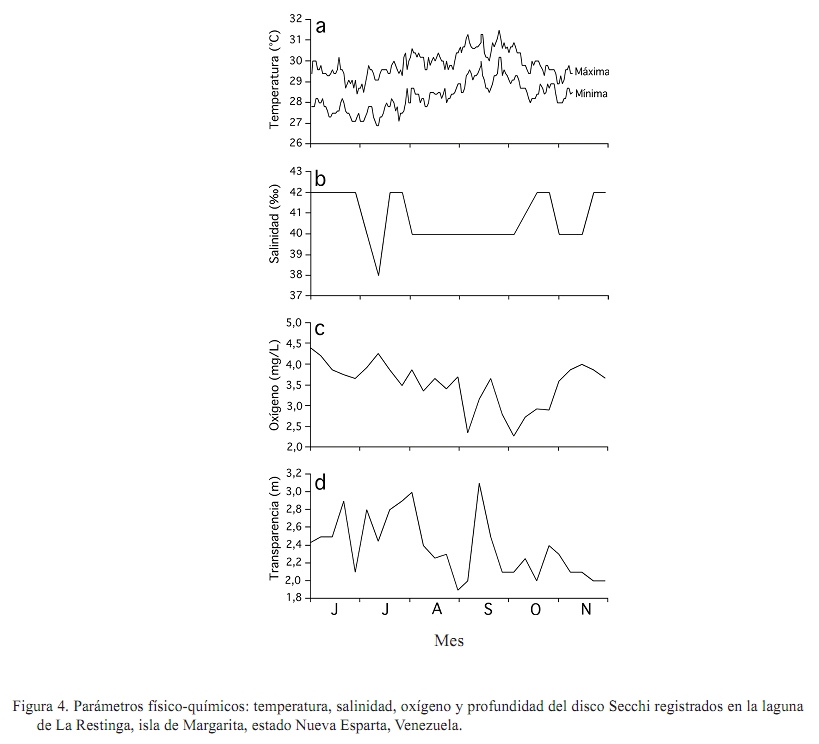
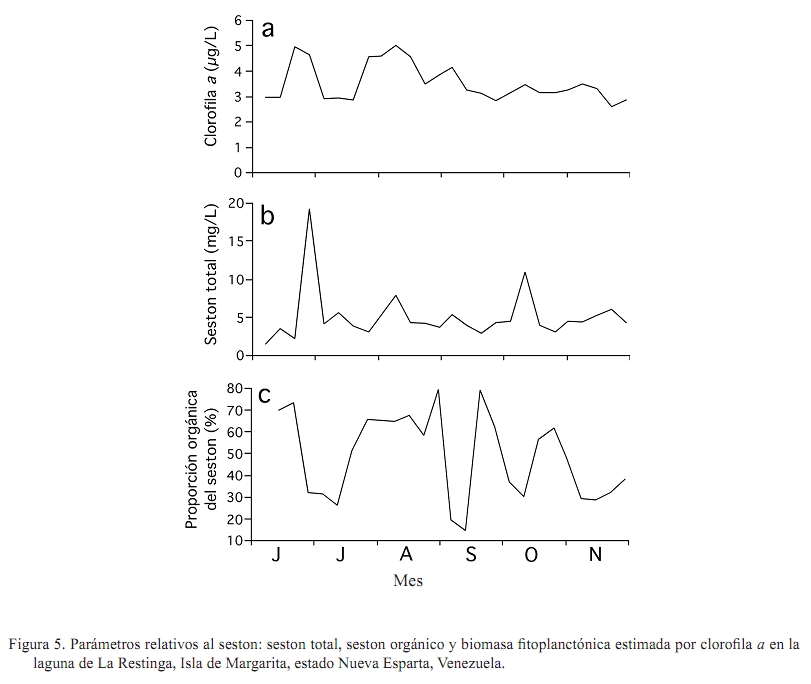
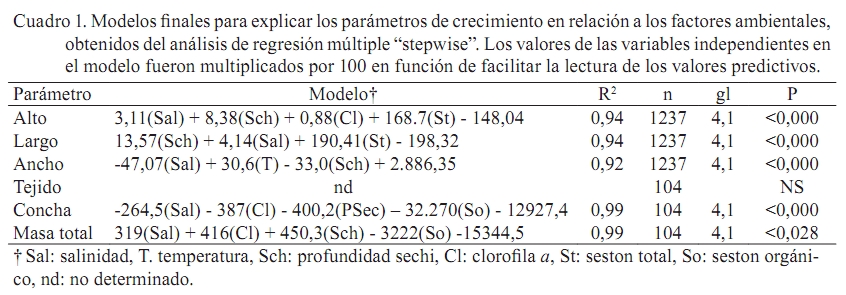
Zootecnia Tropical, Vol. 27, No. 1, 2009, pp. 79-90 Artículos Científicos Identificación de factores que afectan al crecimiento y la supervivencia de la ostra de mangle, Crassostrea rhizophorae (Guilding, 1828), bajo condiciones de cultivo suspendido en la laguna de La Restinga, Isla de Margarita, Venezuela Identification of environmental factors affecting growth and survival of the mangrove oyster, Crassostrea rhizophorae (Guilding, 1828), in suspended culture at La Restinga lagoon, Margarita Island, Venezuela Esperanza Buitrago1, Joaquín Buitrago2, Luis Freites3 y César Lodeiros3* 1 Grupo de Investigaciones en Biología de Moluscos, Universidad de Oriente, Núcleo de Sucre. Cumana 6101, Sucre, Recibido: 06/02/2008 Aceptado: 21/01/2009 Code number: zt09010 RESUMEN La ostra de mangle, Crassostrea rhizophorae, es uno de los moluscos con mayores perspectivas para el desarrollo de la acuicultura en el Caribe y costa atlántica tropical. Aunque se han realizado estudios para establecer su cultivo, no se ha determinado la influencia de los factores ambientales en el desarrollo bajo condiciones de cultivo. Individuos de 30 mm fueron cultivados en cestas españolas a 4 m de profundidad en la laguna de La Restinga, Isla de Margarita, Venezuela. Mensualmente se determinaron la longitud y masa de la concha, así como la masa del tejido blando y los organismos y material depositado sobre las valvas (fouling), además de la supervivencia. La temperatura se registró continuamente y semanalmente se observó la transparencia del agua y se determinaron la salinidad, oxígeno, seston (total, orgánico e inorgánico) y la biomasa fitoplanctónica. La longitud dorso-ventral incrementó 89%, alcanzando tallas de 54,5 mm y 0,48 g de tejido seco, luego de recuperarse de una disminución en los meses de agosto a septiembre (meses cálidos), sugiriendo a la temperatura como un factor importante en el desove. La elevada y constante biomasa fitoplanctónica en el sistema de manglar mostró que la disponibilidad de alimento no fue limitante, lo que en conjunto con la poca variabilidad en los factores ambientales estudiados no permitieron una clara identificación de factores que afectan la varianza observada en la masa de los tejidos blandos y la sobrevivencia de C. rhizophorae en condiciones de cultivo suspendido, con la excepción de la incidencia eventual de gasterópodos depredadores del género Cymatium. Palabras clave: bivalvo, cultivo de moluscos, factores ambientales, predadores, sobrevivencia ABSTRACT Mangrove oyster, Crassosterea rhizophorae, is one of the mollusks with greatest perspectives for aquaculture development in the Caribbean and the tropical Atlantic coast. Even though, many studies have been carried out to establish its culture, there has not been established which and how environmental factors influence its growth under culture condition. Spats with a mean size of 30 mm were suspended in “Spanish baskets” at a depth of 4 m during 6 months. The experiment was carried on at La Restinga lagoon, Margarita Island, Venezuela. Each month oysters were measured (ventral-dorsal and antero-posterior), and weighted (dry weight from conch, tissue and fouling). Survivorship was also evaluated. Water temperature was recorded continuously and weekly measurements of Secchi depth, salinity, oxygen, total, organic and inorganic seston, and chlorophyll a were determined. Results show 89% increase in ventral-dorsal size reaching sizes of 54.5 mm. Dry weight of tissue at the end of the study period was 0.48 g after a decrease during the warmer months of august and september, indicating temperature as an important factor controlling spawning. The high and constant phytoplanktonic biomass present in La Restinga lagoon showed that food availability was not a limiting factor, which together with the small variability in the studied environmental variables, made impossible to identify factors controlling growth and survival of Crassostrea rhizophorae in long-line suspended culture. The exception was the occasional predation by Cymatium snails. Keywords: mangrove oyster culture, growth, predation, survival, environmental factors. INTRODUCCION La ostra de mangle, Crassostrea rhizophorae (Guilding, 1828), es una especie hermafrodita protándrica, de talla mediana-grande, que llega ha alcanzar longitudes de 100 mm, y se distribuye en el Caribe y Atlántico suramericano hasta Brasil. La especie habita normalmente en ecosistemas con manglar y salinidades superiores a 30‰, comúnmente se encuentra adherida a las raíces del mangle rojo, Rhizophora mangle, y es explotada comercialmente, siendo la ostra de mayor producción en el Caribe (Lodeiros et al., 1999). C. rhizophorae es uno de los moluscos con mayores perspectivas para el desarrollo de la acuicultura en el Caribe y la costa atlántica tropical. Su cultivo está principalmente desarrollado en Cuba, país que posee una tecnología de cultivo integral y produce cantidades relevantes (1.000-2.500 t/año) (FAO, 2008). De igual manera en Brasil, Colombia y Venezuela se producen cantidades relativamente importantes. aunque no reportadas por la FAO. En Venezuela, C. rhizophorae supone un recurso pesquero con una actividad económicamente importante de la cual dependen gran número de personas, siendo la única entrada de ingresos en varios grupos familiares (Gil y Moreno, 2007) y se muestra como una especie adecuada para establecer cultivos a través de cooperativas u organizaciones comunales en el occidente y oriente del país (Lodeiros y Freites, 2008). No obstante, a pesar que se han realizado varios estudios e intentos comerciales durante los años 70 y 80, y recientes estudios han permitido diseñar sistemas de captación de semilla y cultivo en suspensión (Buitrago et al., 1999, 2000; Buitrago y Alvarado, 2005; Lodeiros et al., 2006), su cultivo comercial no ha sido desarrollado. Entre los factores que contribuyeron al fracaso de los primeros intentos, se reporta la presencia de gran cantidad de organismos competidores y depredadores (Vélez y Lodeiros, 1990). Revisiones sobre el cultivo de moluscos bivalvos en Venezuela concluyen que existe un limitado conocimiento científico relativo a la identificación de factores que permitan establecer estrategias de cultivo para una producción sostenida (Vélez y Lodeiros, 1990; Vélez, 1991; Lodeiros y Freites, 2008). Las ostras del género Crassostrea, en general, se consideran organismos con capacidad de resistencia a la variabilidad de factores ambientales y son considerados organismos eurihalinos (Quayle, 1988). La respuesta de las ostras, en términos de crecimiento y supervivencia, a las condiciones ambientales ha sido estudiada para C. virginica (Hofmann et al., 1992, 1994; Powell et al., 1995; Dekshenieks et al., 2000) y para C. gigas (Kobayashi et al., 1997; Hyun et al., 2001), destacando el efecto de la disponibilidad de alimento. En C. rhizophorae estas respuestas en condiciones ambientales naturales han sido poco estudiadas. Por otro lado, en un estudio realizado bajo condiciones de cultivo suspendido en el Golfo de Cariaco, durante el periodo de surgencia discontinua y estratificación de la columna de agua (Villarroel et al., 2004) mostraron que el crecimiento C. rhizophorae tan solo alcanzó 15 mm en 6 meses de cultivo, identificando a la disponibilidad de alimento como el factor limitante para el crecimiento y consideraron que tanto la incidencia del “fouling”, en las cestas y sobre la concha, así como la depredación de gasterópodos del género Cymatium fueron factores negativos para el cultivo. El presente estudio evalúa la influencia de factores bióticos y abióticos en el crecimiento y la supervivencia de C. rhizophorae, bajo condiciones de cultivo suspendido en la Laguna La Restinga, Isla de Margarita, Venezuela. MATERIALES Y MÉTODOS El estudio se llevó a cabo durante 6 meses (01 junio al 29 noviembre 2002) en la Laguna de La Restinga, Isla de Margarita, Venezuela (10°, 9874521´ N; 64°, 1621834´ O), con semillas captadas en colectores artificiales construidos con botellas plásticas, según Buitrago et al. (1999). Para el estudio se utilizaron 800 ejemplares de C. rhizophorae de 29,8 ± 5,12 mm (DE) de longitud dorso-ventral de la concha, 1,11 ± 0,06 g de masa de la concha y 0,97 ± 0,05 g de tejido. Los ejemplares se distribuyeron en 8 cestas españolas con una densidad de 100 individuos por cesta, de las que 3 eran réplicas experimentales y 5 de sustitución. Las cestas se suspendieron a 1,5 m de la superficie en una zona de 4 m de profundidad, en una línea larga o “long line” que fue establecido a unos 20 m del manglar, en la zona central de la laguna. Para estimar el crecimiento, mensualmente se determinó la dimensión de la concha en su eje máximo dorso-ventral (largo), antero-posterior (alto) e intervalvar (ancho) en todos los organismos de cada cesta con la ayuda de un vernier digital Mytutoyo (0,01 mm de apreciación). También se obtuvieron quince ejemplares de cada réplica para determinar la masa seca de la concha y tejidos blandos (60°C durante 72 h). De igual manera, fue determinada la masa seca de los organismos y material adherido a la concha o “fouling”. La supervivencia se determinó mensualmente contando la totalidad de los organismos, utilizando los individuos de las réplicas de sustitución para mantener la densidad de 100 individuos por cesta, en las tres réplicas de experimentación. Con el fin de estudiar el efecto de los factores ambientales se determinó la temperatura del agua, oxígeno disuelto y salinidad. Además se evaluó la disponibilidad de alimento a través del seston (total, orgánico e inorgánico), transparencia del agua mediante un disco sechi y biomasa fitoplanctónica, estimada por la determinación de clorofila a. La temperatura se registró a intervalos de 30 min con un termógrafo electrónico (Sealog, Vemco Ltd, Halifax), colocado a la misma profundidad del cultivo. Con una periodicidad semanal se tomaron muestras de agua con una botella Niskin de 2 L de capacidad, a la misma profundidad de las cestas: Luego las muestras fueron transferidas a un contenedor opaco y transportadas al laboratorio en menos de 1 h, cuando fueron procesadas. Submuestras de agua de 1 L, previamente tamizadas con una malla de 153 µm de diámetro de poro fueron concentradas en filtros MSF (0,7 µm de diámetro poro), utilizando un equipo Millipore de filtración al vacío. Los filtros con las muestras concentradas fueron lavados con formiato de amonio al 3% y almacenados a -20°C hasta su posterior análisis. El seston total, inorgánico y orgánico se determinó por métodos gravimétricos y la clorofila a mediante el método colorimétrico según recomendaciones en Stricklan y Pearson (1972). Para la evaluación de los parámetros de crecimiento y la masa seca del “fouling”, se determinó la normalidad de los datos y se aplicó un análisis de varianza de una vía, tomando en cuenta los muestreos realizados en cada mes como tratamientos. Como todos los parámetros mostraron diferencias significativas entre muestreos cuando se les aplicó el ANOVA (P<0,05), se utilizó un análisis “a posteriori” de Duncan (α=0,05), para comparar las medias de los valores absolutos entre los meses sucesivos, en virtud de determinar crecimiento o decrecimiento significativo. Para evaluar el efecto de los factores ambientales en el crecimiento, los valores transformados a log de la tasa diaria específica de crecimiento (división del incremento entre muestreos contiguos y dividido por el número de días de dicho periodo) de las dimensiones de concha, su peso y el de los tejidos de las ostras se contrastaron como variables dependientes con los factores ambientales (variables independientes). Para ello se utilizó un análisis de regresión múltiple “stepwise” usando el programa Statistica (StatSof, 1999). Todas las variables ambiéntales (excepto el fouling) se utilizaron para el análisis de regresión múltiple y mostraron un grado de asociación intermedio-bajo (r<0,7; correlacción de Pearson), por lo que los efectos de colinearidad dentro de los modelos construidos fueron considerados mínimos (Hair et al., 1992). RESULTADOS Crecimiento en longitud y masa El crecimiento en longitud de la concha mostró dos fases bien marcadas para todas las dimensiones (Figura 1a). Una fase de crecimiento acelerado en los dos primeros meses (excepto la longitud entre valvas que fue solo el primer mes) con incrementos significativos (P<0,05), seguida de una fase donde el incremento de la longitud no mostró incrementos significativos (P>0,1). Al final de la experiencia, las ostras incrementaron en su longitud dorso-ventral, alto y entre valvas en un 82,9; 73,5 y 162,7% con respecto a la talla inicial, respectivamente, obteniendo un valor medio de 54,5 ± 9,8 mm de largo, con un intervalo de tallas de 40,5 a 78,1 mm. La masa seca de la concha mostró incrementos significativos (P<0,05) durante todo el periodo de estudio, excepto en el mes de septiembre (Figura 1b). Al final del experimento, la concha alcanzó valores de 14,3 ± 0,79 g. La masa seca de los tejidos de la ostra presentó un crecimiento acelerado hasta finales de julio, cuando alcanzó valores de 0,45 ± 0,042 g (Figura 1c), para luego disminuir su masa en un 49 y 51% durante los meses de agosto y septiembre (0,22 ± 0,032), cuando se observó poco tejido reproductivo, sugiriendo una actividad de desove durante estos meses. Posteriormente, la masa de los tejidos incrementó con tasas similares a las iniciales hasta el final del experimento, cuando alcanzó 0,48 ± 0,057 g, recuperando prácticamente la masa perdida en el período anterior (agosto-septiembre). Supervivencia La supervivencia durante el primer mes fue alta (>95%). En los meses restantes la supervivencia estuvo por debajo del 85% (Figura 2). En el mes de agosto, se encontró una gran dispersión de la media de supervivencia relacionada con la presencia del gasterópodo Cymatium poulseni y el decápodo Callinectes ornatus, lo cual afectó la supervivencia de una de las cestas. Al final del estudio, la supervivencia acumulada fue 53%. “Fouling” en la concha La masa seca del “fouling” sobre las conchas incrementó hasta finales del mes de julio con un valor de 1,67 ± 0,24 g, representando 30% de la masa total de la concha. A partir de esa fecha el “fouling” disminuyó significativamente (P<0,05) hasta el mes de octubre (0,62 ± 0,11 g; 8,3% del peso de la concha), para luego aumentar a 1,29 ± 0,19 g (9% de la concha) en el último mes del experimento (Figura 3). Factores ambientales Durante todo el experimento, las temperaturas se mantuvieron siempre sobre 27°C, con un máximo de 31,6°C (Figura 4a). Entre el periodo agosto-septiembre, la temperatura fue incrementando gradualmente hasta alcanzar su máximo a finales de septiembre (31,3°C), para luego disminuir hasta el final del estudio, cuando fueron registradas temperaturas entre 28 y 29,5°C. Similares valores de temperaturas se observaron a mitad de julio. El rango de temperatura interdiaria se mantuvo generalmente con valores menores a 2°C. En cuanto a la salinidad, ésta varió en unas 4‰ (38-42‰), generalmente se mantuvo entre 4042‰, excepto en el mes de julio, cuando disminuyó a 38‰ debido a fuertes precipitaciones (Figura 4b). El oxígeno disuelto se mantuvo alrededor de 3,5 mg/L, presentando mínimos de 2,27 mg/L en los meses de septiembre y octubre y máximos de 4,5 mg/L a principios del experimento (Figura 4c). Por otro lado, la transparencia osciló entre 1,9 y 3,1 m, manteniéndose generalmente en 2,3 m (Figura 4d), mientras que, la clorofila a se mantuvo generalmente sobre 3 µg/L, con periodos de valores elevados (sobre el rango de 3,5-5 µg/L a mitad de junio-julio) y de finales de julio a mediados de septiembre (Figura 5a). Por último, el seston total mostró valores generalmente de 5 mg/L con maximos a finales de julio (19 mg/L) y la primera semana de octubre (11 mg/L). El porcentaje del seston orgánico se mantuvo variable entre el 2070%, manteniéndose en valores por encima del 60% en el mes de julio, agosto y finales de septiembre. Relación de las tasas de crecimiento y factores ambientales Todos los factores ambientales determinados en el experimento fueron considerados en el análisis múltiple, siendo la salinidad, la transparencia (profundidad del disco secchi), la fracción orgánica del seston y la clorofila a los factores más frecuentemente integrados en los modelos para explicar la variabilidad de los parámetros de crecimiento (Cuadro 1). La integración de todos estos factores explicó más del 90% de la variabilidad de los parámetros de crecimiento, exceptuando la masa del tejido para la cual no pudo construirse un modelo significativo. La salinidad fue el factor predictivo principal en los modelos y la temperatura solo intervino como factor para explicar el ancho de la concha. Por su parte, la clorofila a fue un factor importante para explicar la masa de la concha y la masa total de las ostras. DISCUSIÓN La Laguna de la Restinga es un sistema manglar cerrado que disminuye el efecto que produce la actividad periódica de los vientos alisios que generan surgencia y estratificación de la columna de agua. De esta manera, cambios de los factores ambientales como la temperatura se minimizan debido a la protección que ofrece los manglares (Monente, 1978). Nuestro estudio confirma dicha observación, ya que a pesar que la temperatura posee cierta variabilidad temporal, ésta siempre mantuvo una diferencia de solo 2°C, tanto del rango de temperatura diaria como de todo el periodo de estudio, asemejándose al comportamiento normal de poca variabilidad de zonas tropicales sin influencia de vientos. De igual manera, los otros parámetros físico-químicos como la salinidad, transparencia y oxígeno, mostraron poca variación y a pesar que en nuestro análisis de regresiones múltiples éstos factores ambientales, con excepción del oxígeno, fueron incluidos como variables principales predictivas en los modelos, los resultados indican que las variaciones acontecidas no mostraron una magnitud adecuada como para sugerir cambios en los procesos fisiológicos de moluscos bivalvos (Bernard, 1983; Griffiths y Griffiths, 1987). Por otra parte, una serie de experimentos en laboratorio con C. rhizophorae sugiere que la especie posee una gran capacidad de adaptación a las variaciones de los parámetros físico-químicos como salinidad, temperatura y pH, mostrándose como una especie fuertemente eurioica y resistente a rangos de elevada salinidad y temperatura (Poza y Rodríguez 1987). Esto explica porque la influencia de los factores ambientales en las ostras en la Laguna de La Restinga no fue notable. La única correlatividad observada con la biomasa seca de los tejidos de la ostra fue una disminución significativa en los meses de agosto y septiembre, coincidiendo con el inicio de la elevación de la temperatura del agua característica de estos meses en la zona. Esto podría disminuyendo así la masa seca del tejido durante los meses más cálidos de agosto y septiembre, para empezar la recuperación de la masa de su tejido blando cuando las temperaturas bajaron. Todo esto sugiere que la temperatura podría ser un factor importante que modula la gametogénesis y particularmente el desove de la especie en la zona de estudio. Estudios periódicos e histológicos en los tejidos de Crassostrea rhizophorae en relación a la temperatura y otros factores ambientales son necesarios para contrastar la hipótesis antes señalada. En concordancia con la mayoría de los trabajos de influencia de factores ambientales en el crecimiento de moluscos bivalvos, Powell et al. (1995) y Hyun et al. (2001) afirman que los principales factores que explican el crecimiento de las ostras de las especies C. virginica y C. gigas son la temperatura y la disponibilidad de alimento. De la misma manera, Villarroel et al. (2004) sugirieron que el factor principal del crecimiento de C. rhizophorae en el Golfo de Cariaco, un área influenciada por la surgencia costera, es la disponibilidad de alimento, llegándose a una variabilidad de clorofila a de elevada magnitud (0,2 a 10,5 µg/L), muy diferente a la variabilidad en nuestro estudio, donde los valores se muestran siempre entre 3 y 5 mg/L, los cuales son considerados suficientes para satisfacer las demandas de los moluscos filtradores. De esta manera, en la laguna La Restinga, la disponibilidad de alimento relativo a la biomasa fitoplanctónica no parece ser limitante. A pesar que la clorofila a es una de las variables que se incluyeron en el modelo para explicar la variabilidad de la masa de la concha, su carácter secundario y valor| negativo muestra que no es limitante y que la variabilidad puede ser explicada mas por otros factores ambientales o factores endógenos, como la formación de tejido gonadal que puede exigir energía que podría ser dirigida a los procesos de crecimiento somáticos, tal y como ocurre en bivalvos por razones evolutivas (Toumi et al., 1983). Tal canalización energética preferencial de reservas para la gametogénesis y que afecta el crecimiento somático ha sido señalada para muchos bivalvos, incluyendo los cultivados en el nororiente de Venezuela (Lodeiros y Himmelman, 2000; Narváez et al., 2002; Freites et al., 2003). La ausencia de modelos significativos que pudieran explicar la variabilidad de los tejidos de C. rhizophorae, a través de los factores ambientales estudiados, sugiere la posibilidad de una mayor importancia de la modulación endógena que de la exógena del crecimiento de las ostras bajo cultivo suspendido, en la Laguna de la Restinga. La presencia de organismos epibiontes y material depositado sobre las cestas de cultivo y las conchas (“fouling”) ha sido identificada como una de las principales causas que inhiben y reducen el crecimiento de las ostras (Angell, 1973; Vélez y Lodeiros, 1990; Nascimento, 1991). El “fouling” sobre la concha puede interferir mecánicamente en la acción de apertura de la concha, sea por interferencia en los bordes de la concha o por su peso, el cual genera una fuerza antagónica a la ejercida por el gozne o ligamento. Esto no permitiría abrir la concha cuando el músculo abductor se relaja, afectando a su vez el proceso de filtración (Lodeiros, 2002). No obstante, Lodeiros et al. (2002) en un estudio sobre el efecto de la posición y peso del “fouling” en la concha de C. rhizophorae bajo condiciones de cultivo suspendido, concluyen que es improbable que el “fouling” pueda tener efecto mediante el peso en condiciones de cultivo, por lo que su efecto mas bien podría ser debido a otro tipo de acción ejercida por los epibiontes (esponjas, hidroides, anémonas, cirrípedos, briozoos, tunicados, algas y algunos pocos mejillones (Perna viridis) y particularmente incrustantes como poliquetos espionidos del género Polydora, los cuales pueden afectar el crecimiento de ostras (competidores por el alimento), o bien su aceptabilidad por parte del consumidor (Handley y Bergquist, 1997). El “fouling” sobre las ostras, en el presente estudio, alcanzo 30% de la masa seca de la concha y si bien este valor no es excesivamente alto, se observo que el “fouling” sobre las cestas de cultivo fue bastante alto, aunque no fue evaluado en este estudio. La incidencia del decápodo Callinectes ornatos y particularmente del gasterópodo Cymatium poulseni en una de las cestas experimentales en el mes de agosto condujo a una disminución elevada de la supervivencia, lo cual identifica a la depredación como un importante factor en el cultivo, tal y como lo sugirió Villarroel et al. (2004), en el Golfo de Cariaco, donde existe una mayor incidencia del gasterópodo en los cultivos. Las especies del gasterópodo Cymatium son consideradas uno de los principales factores que afectan de manera negativa al cultivo de moluscos bivalvos en el nororiente de Venezuela (Lodeiros y Freites, 2008) y probablemente del Caribe y costa atlántica, debido a su distribución. En este sentido, Freites et al. (2000) mostraron que en poco tiempo, probablemente un mes, un solo individuo puede acabar con la población incluida en las cestas de cultivo, por lo que se recomienda, al menos en los primeros meses de cultivo, una inspección y retiro continuo de estos organismos dentro de las cestas. También recomienda la realización de estudios de biocontrol con organismos como erizos que puedan eliminar larvas y organismos juveniles de este gasterópodo. Una alternativa para la eliminación de depredadores y organismos del “fouling” podría ser el sometimiento de las cestas de cultivo a desecación por un período prolongado, tal como ocurre, debido a las mareas, con las ostras en condiciones naturales en el manglar; de esta manera, el cultivo en la zona intermareal podría ser una estrategia adecuada para C. rhizophorae. En cuanto al crecimiento se refiere, las ostras cultivadas obtuvieron un incremento de 24,7 mm en 6 meses, el cual representó 83% de la dimensión inicial de la concha con una supervivencia acumula del 53%. Este crecimiento fue alto, comparado con ostras provenientes de la misma cohorte y cultivadas en Turpialito, Golfo de Cariaco (44,6%), bajo las mismas condiciones (Villarroel et al., 2004). En contraste, el mismo crecimiento fue bajo si los comparamos con los obtenidos, también en 6 meses, por Buitriago et al. (2000), en la misma zona de estudio, donde el incremento de la concha fue 128%, mientras que el observado en Cubagua por Hernández et al. (1998) fue 189%. A diferencia del presente estudio, en los dos estudios antes mencionados los ejemplares fueron cultivados en linternas que fueron suspendidas de balsas flotantes. Por otra parte, Angell (1973) en un estudio de cultivo de C. rhizophorae, en la Laguna de La Restinga, fijadas en colectores de asbesto y suspendidas en balsa encontró mayor crecimiento que en todos los estudios anteriores (68 mm en 6 meses); sin embargo, la supervivencia fue 10%, imputando este resultado a la baja densidad de organismos por área. Lodeiros et al. (2006) en un estudio realizado para seleccionar el tipo de cesta y sistema de cultivo (“long line” vs. balsa), en la misma zona de estudio, determinó un mayor crecimiento en balsa y con cestas tipo linternas. La diferencia de estos estudios radica en la utilización de sistemas (balsas y “long line”) y recintos de cultivo diferentes (cestas, asbesto) con diferente densidad de siembra, lo cual sugiere la realización de estudios multifactoriales que contemplen diferentes densidades, diseños de cestas y sistemas de cultivo. Al final de la experiencia, las ostras incrementaron entre valvas en 163% con respecto a los valores iniciales, a diferencia de las dimensiones dorso ventral y antero-posterior de la concha (<83%). Este incremento entre valvas del doble de las otras dimensiones generó una morfología particular en la ostra, donde la valva izquierda se muestra de forma cóncava y la derecha y superior plana, generando mayor cavidad intervalval y con ello mayor biomasa que las encontradas en la naturaleza de la misma talla dorso-ventral y antero-posterior (observación personal). Esta característica de las ostras cultivadas en suspensión le confiere una mayor producción de la vianda y parece estar asociada al método de cultivo sin fijación en un sustrato, como ocurre en la naturaleza. Estudios comparativos de estas dos situaciones con análisis morfo y gravimétricos son recomendados. CONCLUSION Nuestros resultados muestran poca variabilidad ambiental en el sistema de manglar de la Laguna de la Restinga unido con una elevada biomasa fitoplanctónica y por ende la inexistencia de identificación de factores ambientales que afecten de manera significativa la varianza observada en la masa de los tejidos blandos y la sobrevivencia de C. rhizophorae, en condiciones de cultivo suspendido en long line. AGRADECIMIENTOS Esta investigación se llevó a cabo cofinanciamiento del proyecto FONACIT N° 2000001415 titulado “Transferencia tecnológica del cultivo de la ostra de mangle, Crassostrea rhizophorae en el estado Nueva Esparta. Agenda Pesca y Acuicultura” Se agradece la asistencia técnica de Karem Lunar, Pulido Moreno, Zoila Vázquez y Juan Carlos Capelo. La Fundación para la Investigación y Desarrollo de la Acuicultura del estado Sucre contribuyó parcialmente en esta investigación, siendo ésta su contribución No. 9. LITERATURA CITADA
Copyright © 2009 - Zootecnia Tropical The following images related to this document are available:Photo images[zt09010f5.jpg] [zt09010f4.jpg] [zt09010f1.jpg] [zt09010f3.jpg] [zt09010f2.jpg] [zt09010c1.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}