 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
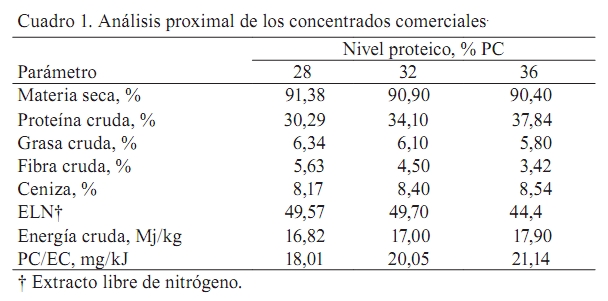
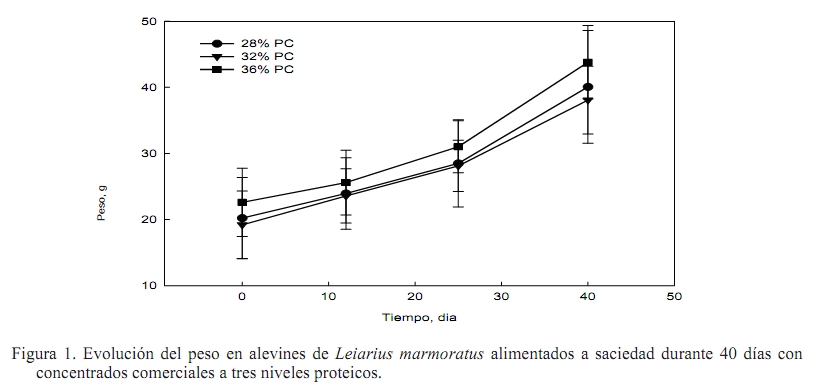
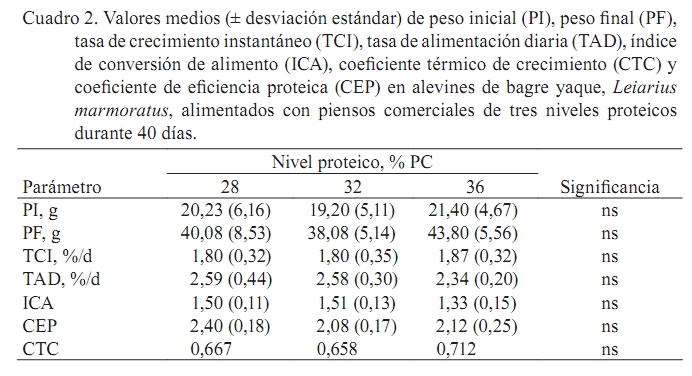
Zootecnia Tropical, Vol. 27, No. 2, 2009, pp. 187-194 Artículos Científicos Influencia del contenido proteico en el crecimiento de alevines de bagre yaque, Leiarius marmoratus, alimentados con concentrados comerciales Effect of dietary protein level on growth performance of fingerlings of catfish yaque, Leiarius marmoratus, fed with three commercial diets José A. Mora Sánchez1*, Francis Moyetones1 y Miguel Jover Cerdá2 1Estación de Piscicultura, Decanato de Agronomía, Universidad Centroccidental “Lisandro Alvarado” Apartado Postal 400, Recibido: 20/06/08 Aceptado: 27/11/08 Code number: zt09021 RESUMEN Se realizó un ensayo de alimentación para evaluar el efecto de 28, 32 y 36% de proteína cruda en concentrados comerciales extrusionados sobre el crecimiento de alevines de bagre yaque, Leiarius marmoratus. Un total de 180 alevines (20,3 g) fueron dispuestos en 12 jaulas (0,5 m3), con cuatro réplicas por tratamiento. Las jaulas se distribuyeron al azar en cuatro tanques (2,0 m3), a razón de tres jaulas/tanque. Cada tanque se dotó con flujo continuo de agua (1,5 L/min), aireación y drenajes independientes. Los concentrados fueron administrados a saciedad aparente durante 40 días. No se encontraron diferencias significativas en la tasa de crecimiento instantáneo (TCI), tasa de alimentación diaria (TAD), índice de conversión del alimento (ICA), coeficiente térmico de crecimiento (CTC) y coeficiente de eficiencia proteica (CEP) en los alevines alimentados con concentrados de diferente nivel proteico. La TCI correspondió a 1,80%/d. La TAD fluctuó de 2,34 a 2,6% PV/d, el ICA entre 1,33 y 1,5, el CTC entre 0,667 y 0,712 y el CEP varió de 2,08 a 2,4. Los niveles proteicos estudiados tuvieron el mismo efecto sobre el crecimiento, conversión del alimento, tasa de alimentación diaria, coeficiente térmico de crecimiento y coeficiente de eficiencia proteica en alevines de Leiarius marmoratus. Palabras clave: Yaque, Leiarius marmoratus, bagres, Pimelodidae, coeficiente térmico. ABSTRACT An experiment was conducted to evaluate the effect of dietary protein level on growth performance of bagre yaque, Leiarius marmoratus, fingerlings fed with three commercial extruded diets. Fish (20.3 g) were placed in twelve 0.5 m3 cages at a density of 15 fish per cage. Groups of three cages were placed inside each of four 2 m3 interior tanks. Three commercial diets or treatments (28, 32, and 36% crude protein) were randomly assigned to cages inside each interior tank to adjust to a completely randomized block design with four replicates. Each interior tank was provided with continuous water flow (1.5 L/min), aeration and independent drains. Diets were provided at apparent satiety for 40 days. At the end of the experiment there were no significant differences (P >0.05) on specific growth rate (SGR), daily feed consumption (DFC), daily feed conversion ratio (FCR), thermal growth coefficient (TGC), and protein efficiency rate (PER) of fingerlings fed the three commercial diets. SGR was 1.8%/d. The DFC varied between 2.34 and 2.6% BW/d, FCR was between 1.33 and 1.5, and PER varied between 2.08 and 2.4. The protein levels studied had the same effect on growth, feed conversion rate, daily food, and protein efficiency ratio on fingerlings of Leiarius marmoratus. Keywords: Leiarius marmoratus, catfish, Pimelodidae, thermal growth coefficient, TGC. INTRODUCCION El bagre yaque, Leiarius marmoratus (Gill, 1870) (Siluriforme:Pimelodidae), es una especie que se distribuye en las cuencas de los ríos Orinoco y Amazonas (Castillo, 2001; Ramírez y Ajiaco, 1997). En ambientes naturales presenta hábitos alimentarios piscívoros (Layman et al., 2005); sin embargo, algunos autores destacan una tendencia omnívora (Ramírez y Ajiaco, 1997; Novoa, 2002). Esta especie presenta cualidades para ser incorporada a la piscicultura dada la excelente calidad de su carne blanca, la cual posee pocas espinas, textura y sabor de apreciable valor gastronómico. A partir de 1986 se practica su reproducción inducida (Kossowski, 1986; Escobar y Mojica, 1997), lo cual ha permitido la sistematización de la producción de alevines a pequeña escala, sustentada en reproductores de segunda y tercera generación (Mora, 2003; Mora y Kossowski, 2006). Además, se han realizado experiencias en la obtención de híbridos con bagre rayado, Pseudoplatystoma fasciatum y Pimelodus blochii (Kossowski, 1991, 1992a,b, 1994, 1996a,b). Sin embargo, es necesario avanzar en la información referida a sus necesidades nutricionales y su alimentación, considerando la alta incidencia de este rubro en los costos de producción en sistemas acuícolas. En este sentido, se ha constatado que a partir de la reabsorción del saco vitelino (48 h a 27ºC) se presenta una fase de canibalismo que puede ser superada con suministro de alimento vivo (nauplios de Artemia) y luego subsecuentemente, en fases avanzadas, por alimentos húmedos preparados a base de hígado fresco de pollo (licuado), mezcla de hígado licuado y alimentos (molido) para peces, hasta lograr la aceptación de concentrados comerciales extrusionados a los 30 días de edad. Desde este momento, es factible realizar su pre-engorde y engorde con el uso exclusivo de alimentos comerciales (Kossowski, 1992b). No obstante, los alimentos comerciales disponibles en el mercado nacional están formulados para engorde de tilapias, cachamas y sus híbridos en estanques de tierra. Así, Silva Acuña y Guevara (2002) evaluaron el crecimiento en juveniles de híbridos de cachamas en estanques utilizando alimentos comerciales de 24 y 28% proteína cruda, sin encontrar diferencias significativas a nivel de los pesos finales. No obstante, información de esta índole es inexistente en Leiarius marmoratus. Por tal razón, el objetivo del presente trabajo fue evaluar el efecto de la concentración de diferentes niveles proteicos en alimentos comerciales sobre el crecimiento, tasa de alimentación, conversión del alimento y coeficiente de eficiencia proteica en alevines de bagre yaque, L. marmoratus. MATERIALES Y METODOS El ensayo se realizó en la Estación de Piscicultura, adscrita al Decanato de Agronomía de la Universidad Centroccidental “Lisandro Alvarado” , en el Municipio Peña (Yaracuy, Venezuela). Procedencia de los peces Los alevines fueron producidos en mayo 2006 mediante reproducción inducida aplicando una dosis de 6 µg de LHRHa/kg de hembra (Mora y Kossowski, 2006). En los primeros meses de edad, los alevines fueron alimentados con un concentrado comercial para tilapias (28% proteína cruda, PC y 3% de grasa cruda, GC). En septiembre 2006, 180 alevines (20,3 ±1,2 g) fueron seleccionados, trasladados a los tanques de experimentación y dispuestos según el diseño experimental planteado. Alimentos y aplicación Se utilizaron dos alimentos comerciales (Vitalim C.A., Chivacoa, Venezuela), los cuales son extrusionados para peces, con 28 y 36% de PC y 6,34 y 5,8% de GC, respectivamente. Estos se aplicaron en tres esquemas de alimentación: Un primer grupo de peces recibió el alimento con 28% PC (T1), el segundo grupo una mezcla a partes iguales (50:50) de ambos alimentos para conseguir un contenido proteico de 32% PC (T2) y un tercer grupo, el alimento con 36 % PC (T3). Con el propósito de adecuar el tamaño del alimento a la boca de los alevines, los concentrados fueron molidos, doblemente cernidos, y finalmente seleccionadas partículas homogéneas de 1,2 mm Ø. Se realizó el análisis proximal (AOAC, 1990) a muestras de estos alimentos, determinando humedad por secado en estufa a 110ºC hasta peso constante. El contenido de cenizas se determino por incineración de las muestras en una mufla a 500ºC hasta peso constante (AOAC, 1990). El contenido de proteínas fue calculado a partir del contenido de nitrógeno total de muestras analizadas por el método Kjedahl. Los lípidos totales fueron determinados por extracción en frío con una mezcla de cloroformo/metanol (Folch et al., 1957) y la fibra por digestión ácido detergente. La energía bruta se determinó mediante un calorímetro adiabático. Los valores resultantes del análisis se presentan en el Cuadro 1. Diseño experimental Los alevines se distribuyeron en tres grupos de peces y en cada uno se dispusieron cuatro réplicas. Para ello fueron asignadas 12 jaulas (0,5 m3), confeccionadas en malla plástica, destinando cuatro unidades por grupo con 15 peces/jaula. Las jaulas se distribuyeron al azar en cuatro tanques (2,0 x 1,0 x 1,0 m), a razón de tres jaulas/tanque. Cada tanque se dotó con flujo continuo de agua (1,5 L/min), aireación y drenajes independientes. A fin de evitar el eventual desplazamiento del alimento entre jaulas, las mismas fueron individualmente provistas de un comedero central, constituido por un cilindro plástico de 30 cm Ø y 30 cm de alto. La alimentación se realizó a saciedad aparente y se estableció mediante la aplicación de pequeñas cantidades de alimento durante un período continuo de 30 min, registrándose la ingesta total como nivel de saciedad aparente. El alimento se suministró dos veces al día, mañana (9:30-10:00 h) y tarde (15:30 – 16:00 h) durante 40 días continuos. Los peces fueron aclimatados por 15 días, y en ese lapso alimentados a saciedad con el concentrado comercial de 28% PC. Registros biométricos Se realizaron muestreos biométricos a 12, 25 y 40 días en la totalidad de los peces de cada jaula. En cada ocasión se obtuvo el peso individual promedio, utilizando una balanza electrónica (Mettler 2000, Suiza) con precisión de 0,1 g. Posteriormente, se calcularon los promedios de peso (± desviación estándar) y biomasa (kg) de cada jaula. Al final del ensayo se determinó: Tasa de crecimiento instantáneo (TCI), según Brown (1957), por la expresión: TCI (%/d) = (ln P( t + ∆t ) – ln Pt/t) x 100 Tasa de alimentación diaria (TAD), según Weatherley y Gill (1987), mediante: TAD (%/d)= (alimento consumido (kg)/biomasa media)/t Índice de conversión del alimento (ICA) se calculó como: ICA = alimento consumido (kg)/incremento biomasa (kg) Coeficiente de eficiencia proteica (CEP) se obtuvo por la relación: CEP = incremento de biomasa (kg)/proteína ingerida (kg) Además, se calculó el coeficiente térmico de crecimiento (CTC) propuesto por Iwama y Tautz (1981) y discutido por Cho y Bureau (1998) y Jobling (2003), según: CTC = (Pf 0,3333 – Po 0,3333)/Σ (T x d) x 100 donde Pf = peso final, Po = peso inicial, T = temperatura del agua (ºC) y d = días. En los tanques se determinó diariamente, antes de la aplicación del alimento, los niveles de oxígeno disuelto, utilizando un oxímetro (YSI-57, Yellow Springs, Ohio, EUA), pH y nitritos. La temperatura del agua se mantuvo constante (26 - 27ºC) mediante la utilización de resistencias eléctricas dispuestas en el tanque-reservorio de distribución de agua al sistema. Los niveles de oxígeno disuelto se mantuvieron por encima de 5 mg/L. Análisis estadístico Los valores de peso final (PF), TCI, TAD, ICA y CEP fueron sometidos al análisis de varianza de una vía, en función del nivel proteico (Steel et al., 1997). En todos los casos se utilizó el peso inicial como covariable. En los análisis se adoptó un nivel de significancia de 5%. En el procesamiento de datos se utilizó el programa estadístico Statgraphic Plus (Statpoint, 2005). RESULTADOS No se presentó mortalidad en ningún tratamiento y los peces exhibieron una condición saludable al final del ensayo. La evolución de los pesos promedios por tipo de alimento se presenta en la Figura 1. No se encontraron diferencias significativas (P>0,05) en ninguno de los parámetros estudiados (Cuadro 2). El peso final varió entre 38,08 g (32% PC) y 43,8 g (36% PC). Los promedios de TCI fluctuaron entre 1,8 (28 y 32% PC) y 1,87 %/d (36% PC) y la TAD correspondió a 2,34 (36% PC) y 2,59% biomasa/d (28% PC). El ICA fue 1,33 para el alimento con 36% PC y no presentó diferencias estadísticamente significativas (P>0,05) respecto al promedio de 1,50 obtenido con 28 y 32% PC. El coeficiente térmico de crecimiento no presentó diferencias estadísticamente significativas y fluctuó entre 0,667 (28% PC) y 0,712 (36% PC). En cuanto al CEP, el valor mínimo fue 2,08 (32% PC) y el máximo 2,40 (28% PC). DISCUSION La comparación de los parámetros de crecimiento de bagre yaque (L. marmoratus) con la información disponible en otras especies de bagres muestra que su crecimiento fue superior al bagre bagrid (Mystus nemurus), similar al bagre de canal (Ictalurus punctatus) e inferior a los bagres jundía (Rhamdia quelen) y pintado (Pseudoplatystoma coruscans). No obstante, el crecimiento de los alevines de bagre yaque en laboratorio fue inferior al obtenido en peces de mayor tamaño en jaulas flotantes donde se obtuvo una TCI de 2,78 %/d (Mora y Jover, 2005). En el presente ensayo se constató que el bagre yaque exhibió avidez y aceptación de los piensos comerciales a las tasas de alimentación del orden de 2,5 %/d, similares a las citadas en el bagre bagrid (Ng et al., 2000) y bagre de canal (Gaylord y Gatlin, 2001), e inferiores a las del bagre jundía (Salhi et al., 2004) y pintado, que mantuvieron mejor crecimiento. Sin embargo, al menos en bagre jundía, a una TAD menor (2,1 %/d) también redujo su crecimiento (Meyer y Fracalossi, 2004). En cuanto al efecto de la concentración de proteína en la dieta, en general se reconoce un aumento en el crecimiento a medida que se incrementa la proteína en el alimento lo cual ocurre hasta un determinado nivel, a partir del cual el crecimiento se reduce. Este efecto se ha señalado para el bagre bagrid (M. nemurus), encontrándose un incremento del crecimiento con piensos de proteína dietaria creciente desde 20 hasta 41% PC, mientras que con 47 y 53% PC el crecimiento se afectó negativamente, por lo cual se estableció un nivel óptimo de 44% PC (Ng et al., 2001). En el bagre jundía (R. quelen), especie omnívora, se ha referido a nivel de crías (0,3 a 1,5 g) un incremento del crecimiento y mejora de la conversión de alimento asociada al incremento del nivel proteico de 30 a 38% PC, pero a niveles proteicos mayores en torno a 43% PC el efecto es negativo y así, se reconoció un nivel óptimo de 37% PC (Salhi et al., 2004). No obstante, en esa especie otros autores destacan un crecimiento similar de alevines de 1,5 g con niveles proteicos de 33 y 37% PC (Meyer y Fracalossi, 2004). Para otro bagre omnívoro, como el bagre de canal (I. punctatus) se observó una mejora del crecimiento con niveles proteicos de 37 frente a 32% PC, asociados a una reducción de la tasa de alimentación (Gaylord y Gatlin, 2001). Estos niveles coinciden con las recomendaciones de Wilson y Moreau (1996), Robinson et al. (2001) y Robinson y Li (2005) señalando 35% PC para alevines de esa especie. El efecto antes descrito no se obtuvo en este ensayo y así, a pesar del incremento del nivel de proteína en el alimento, la TCI no presentó diferencias significativas. Sin embargo, con 36% PC el valor absoluto de la TAD (2,34%/d) y el ICA (1,33) tendieron a disminuir respecto a 32 y 28% PC, aunque estas diferencias resultaron estadísticamente no significativas. Es de resaltar que resultados similares se obtuvieron en el engorde hasta talla comercial de bagre yaque producido en jaulas (Mora y Jover, 2005), lo cual reafirma el carácter omnívoro en la especie. Los altos niveles proteicos referidos en el bagre jundía (Salhi et al., 2004) pueden estar justificados en virtud del tamaño de los peces (0,3 a 1,5 g), puesto que en general los peces en sus etapas tempranas de vida presentan una alta tasa metabólica que a su vez requiere mayor suministro proteico (Lovell, 1989). De acuerdo a ello, los alevines en este ensayo con pesos de 20 g necesitarían un menor nivel de proteína dietaria en respuesta a una menor tasa metabólica. En este sentido, el crecimiento del bagre bagrid (M. nemurus) alimentados con niveles proteicos 29 y 36% PC (Ng et al., 2001), comparables a los usados en este ensayo, resultaron inferiores (0,97 y 0,89%/d) e incluso a niveles proteicos superiores (41 y 47% PC) a pesar del menor tamaño de los alevines (7 g), por lo que es claro que L. marmoratus creció mejor. Por otro lado, también el nivel de lípidos en el pienso tiene un efecto sobre el crecimiento. En esta forma, en bagrid (M. nemurus) el incremento de lípidos en el pienso, de 8 a 14%, mejoró el crecimiento en todos los niveles proteicos (Salhi et al., 2004). En bagres carnívoros, como el pintado (Pseudoplatystoma coruscans), también se ha encontrado que niveles crecientes de inclusión de lípidos, a base de aceite de soya, en el alimento (46% PC) mejoran la tasa de crecimiento, eficiencia proteica y reduce la tasa de alimentación (Martino et al., 2002). Los alimentos con un nivel de lípidos de 6,1% promovieron un menor crecimiento (1,1%/d), mientras que con niveles de lípidos 10; 14,8 y 18,1% se obtuvieron crecimientos de 2,4; 2,8 y 4,1%/d, respectivamente (Martino et al., 2002). En esa especie otros trabajos ratifican que en raciones de 46% PC y con inclusión creciente de lípidos de 10,1 a 18,1%, a base de manteca de cerdo o aceites de linaza o maíz, se obtuvo una mejora significativa de la conversión de alimento e incluso puede alcanzar 0,8 con piensos de 46/18,1 (Campos et al., 2006). El efecto de ahorro de proteínas por inclusión de lípidos también se puede observar en el bagre jundía (R. quelen) al considerar los piensos 30/14 y 34/8, obteniéndose en este último un mejor crecimiento; en los piensos 34/14 y 38/8 que mostraron un mismo crecimiento; y además, en los piensos 38/14 y 43/8 donde el primero presentó mayor TCI. No obstante, este efecto de ahorro de proteína no se apreció en el bagre canal, donde inclusiones de lípidos de 4,5 a 11% de grasa cruda no mejoraron el crecimiento. En esta especie se ha demostrado el efecto positivo de la inclusión de carbohidratos en el ahorro de proteína dietaria (Lovell, 1989; Robinson y Li, 2005). En atención a la información disponible en estas especies de bagres, también cabría esperar que el crecimiento de bagre yaque pueda ser mejorado por la inclusión de mayores niveles de lípidos en el pienso y este aspecto constituye un tema de interés que debe ser considerado para futuros estudios, toda vez que la reducción del nivel proteico en el pienso está asociado a un menor costo del alimento. En conclusión, los niveles proteicos de 28, 32 y 36% PC en los alimentos comerciales estudiados tuvieron el mismo efecto sobre el crecimiento, conversión del alimento, tasa de alimentación diaria, coeficiente térmico de crecimiento y coeficiente de eficiencia proteica sobre alevines de Leiarius marmoratus con pesos de 20 g y alimentados a saciedad aparente. AGRADECIMIENTOS Los autores expresan su agradecimiento al Consejo de Desarrollo Científico, Humanístico y Tecnológico (CDCHT) de la Universidad Centroccidental “Lisandro Alvarado” por el financiamiento parcial de la investigación a través del Proyecto CIAG-052AG2002. De igual forma al personal de la Estación de Piscicultura, en especial a los Señores José Parada y Carlos Apóstol. LITERATURA CITADA
Copyright © 2009 - Zootecnia Tropical The following images related to this document are available:Photo images[zt09021c2.jpg] [zt09021c1.jpg] [zt09021f1.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}