 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
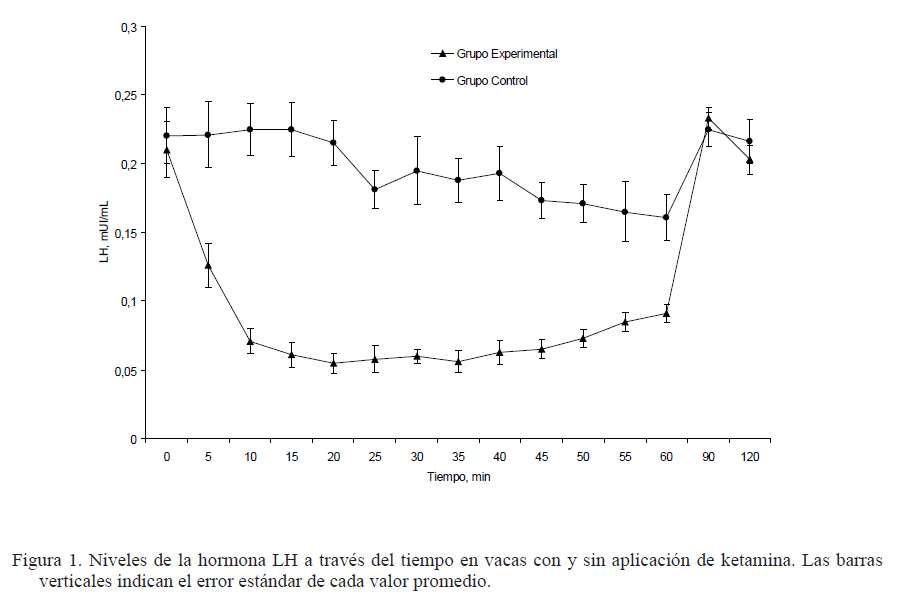
Zootecnia Tropical, Vol. 27, No. 3, 2009, pp. 271-276 Ketamina disminuye los niveles de la hormona luteinizante en vacas lecheras Ketamine decreases the level of luteinizing hormone in dairy cows Iraima Medina y Nelson L. Daló* Universidad Centroccidental Lisandro Alvarado, Decanato de Ciencias Veterinarias, Unidad de Investigación Dr. H. Moussatché. Apartado Postal 722, Barquisimeto, Lara. Venezuela. *Correo electrónico: nelsondalo@yahoo.com Code Number: zt09030 RESUMEN El hipotálamo integra las señales excitatorias mediadas por glutamato que desencadenan en la liberación de la hormona liberadora de gonadotropinas (GnRH), la cual regula la secreción adenohipofisiaria de la hormona luteinizante (LH), quien tiene un papel fundamental en el ciclo reproductivo. Estas acciones neuro endocrinas del glutamato son susceptibles de ser bloqueadas por drogas antagonistas del receptor N-metil-D-aspartato (NMDA), entre ellas, la ketamina. El objetivo de este trabajo fue determinar la disminución en los niveles séricos de la LH en las vacas a dosis bajas de ketamina. Se usaron 12 vacas de la raza Carora, no preñadas y con signos de celo, divididas en dos grupos. Seis de ellas recibieron ketamina (1 mg/kg, IV) diluida en 20 mL de solución salina al 0,85% y seis controles inyectadas con el diluyente. Muestras de sangre fueron tomadas cada 5 min y la LH fue cuantificada por quimioluminiscencia. La ketamina produjo a los 5 min de la inyección niveles de LH significativamente bajos (P<0,001) que se mantuvieron durante 50 min. Después de los 55 hasta los 120 min los valores de la LH retornaron a su nivel control. Este potente efecto de la ketamina sobre la LH nos hace sugerir que situaciones similares ocurrirían con otras sustancias antagonistas que alteren el sistema glutamatérgico, podrían también interrumpir el ciclo reproductivo. Palabras clave: ketamina, hormona luteinizante, LH, GnRH. ABSTRACT The hypothalamus integrates excitatory signals mediated by glutamate that result in the release of gonadotropin releasing hormone (GnRH) that regulates the adenohypophysis luteinizing hormone (LH) secretion, which has a pivotal role in reproductive cycle. This neuro endocrine action of glutamate is susceptible of being blocked by N-methyl-D-aspartate (NMDA) receptor antagonists, among others, ketamine. The main goal of this work was to determine whether a low dose of ketamine induces a decrease in the plasmatic level of LH in cows. Twelve Carora breed dairy cows, not pregnant and showing signs of estrous were used. Cows were divided in two groups. Six of them were injected with ketamine (1 mg/kg, IV), diluted in 20 mL of 0.85% saline solution and six were injected with the solvent. Blood samples were taken every 5 min and the LH levels were quantified by chemiluminescence. Ketamine produced 5 min after its injection a significant decrease in LH level that lasted 50 min. From 55 to 120 min, the LH levels returned to control values. This potent effect of ketamine on the LH let us suggest that similar situation may occur with others antagonistic substances that alter the glutamatergic system, may also disrupt the reproductive cycle. Keywords: ketamine, luteinizing hormone, LH, GnRH. INTRODUCCIÓN El hipotálamo es el centro donde se integran y procesan las informaciones procedentes del propio sistema nervioso central, del exterior (luz, estímulos olfatorios, alimentación) y de las gónadas (hormonas ováricas). De esta integración resultan señales excitatorias mediadas por el glutamato (Van Den Pol y Trombley, 1993; Hrabovszky y Liposits, 2008) y también señales inhibitorias mediadas por el ácido gamma amino butírico (GABA) (Decavel y Van Den Pol, 1990) que desencadenan la liberación de GnRH hipotalámica, la cual estimula la secreción de las gonadotropinas: la hormona luteinizante (LH) y la hormona folículo estimulante (FSH) (Bourguignon et al., 1997). Estas, conjuntamente con las hormonas esteroideas regulan la periodicidad de los ciclos reproductivos (Brann y Mahesh, 1994). El concepto que se tenía que las neuronas hipotalámicas usaban péptidos, neuromoduladores y factores de liberación de acción lenta como sustancias neuroactivas ha cambiando totalmente (Van Den Pol y Trombley, 1993) En el hipotálamo, el glutamato es el principal neurotransmisor excitatorio y realiza su acción por interacción con receptores ionotrópicos y metabotrópicos. Se conocen dos grandes tipos de receptores ionotrópicos: los receptores NMDA, denominados así por el nombre de su principal agonista sintético, el ácido N-metil-D-aspártico (NMDA) y los receptores no-NMDA. Los receptores NMDA median despolarizaciones post-sinápticas lentas y los receptores no-NMDA median despolarizaciones rápidas (Brann y Mahesh, 1994). No existe acuerdo en que tipo de receptores son principalmente activados para inducir la liberación de GnRH. Se han reportado evidencias que indican que durante el proceso de liberación de la GnRH, el glutamato interactúa con los receptores no-NMDA (López et al., 1992) y otros indican que es a través de receptores NMDA (Carbone et al., 1996, Downing et al., 1996) o en ambos (Brann y Mahesh, 1994). Es importante desde el punto de vista fisiológico conocer el mecanismo involucrado en la liberación de LH y, del mismo modo, dilucidar porqué en ciertas circunstancias su patrón de secreción se ve modificado con una alteración de la actividad ovárica. La LH mantiene ciertos niveles durante el ciclo estrual que durante el celo alcanza un pico que determinan la maduración, ruptura del folículo y el proceso de ovulación (Fortune et al., 2004). Se podría hipotetizar que si el sistema glutamatérgico es alterado por factores ambientales, enfermedades o por drogas antagonistas de sus receptores, se disminuirían los niveles hipotalámicos de la GnRH y se desencadenaría una inhibición en la liberación de LH y no ocurriría la ovulación. Existen muchas drogas que pueden inhibir la acción central del glutamato. Las más conocidas son los anestésicos volátiles e intravenosos, los barbitúricos, los alcoholes (Franks, 2008), agentes contaminantes como el uretano (Koblin, 2002), los anticonvulsivos (Piña Crespo y Daló, 2007), antitusígenos como el dextrometorfano y una gran cantidad de drogas que se ensayan como neuroprotectores en enfermedades neurológicas (Kalia et al., 2008). Pero existe la posibilidad de que agentes agregados a los alimentos como el glutamato monosódico o el ácido aspártico puedan aumentar la actividad secretoria neuroendocrina (Urbansky et al., 1994; Downing et al., 1996). Entre las drogas inhibitorias están la ketamina, la tiletamina y la fenciclidina, que forman parte del arsenal terapéutico veterinario y usados eventualmente como pre anestésicos, anestésicos disociativos o agentes inmovilizantes en grandes animales. Estos son drogas antagonistas no competitivos del receptor NMDA que pasan fácilmente la barrera hematoencefálica y producen un bloqueo muy potente del canal iónico (Reynolds y Miller, 1990). Estas evidencias han permitido sugerir que si se desea evitar un ciclo estrual anovulatorio, un agente que bloquee los receptores NMDA no debería ser administrado durante el celo. Este trabajo tuvo como objetivo determinar si dosis bajas de ketamina producen disminución en los niveles de LH. Estos resultados son de utilidad para alertar sobre el correcto uso de estas drogas y cuando se deseen investigar causas exógenas de baja fertilidad, que es el problema de mayor relevancia en el rebaño lechero. MATERIALES Y MÉTODOS Estos experimentos fueron realizados en la estación experimental “Manuel Salvador Yépez”, explotación lechera con un manejo semi intensivo, ubicada el El Torrellero, municipio Simón Planas, estado Lara, Venezuela. Se usaron un total de 12 vacas lecheras de la raza Carora, no preñadas, con un peso promedio de 477 ± 51,46 kg. Del total de vacas seleccionadas, cinco tenían dos partos y siete, tres partos. El último parto ocurrió entre los meses de noviembre a febrero. Los animales fueron alimentados en pastizales de Brachiaria decumbens y B. humidicola y suplementados con alimento concentrado, que recibieron a razón de 3,5 kg/vaca, en promedio. Antes de iniciar el experimento cada animal fue palpado para determinar la presencia de folículo y ausencia de cuerpo lúteo y sólo fueron incluidas las clínicamente sanas en condición de celo. El celo fue considerado positivo cuando fueron observados signos, como presencia de limo claro y filante en la vulva, enrojecimiento e edema vulvar, inquietud, disminución de pastoreo, conductas de montar a otras hembras o dejarse montar (Sepúlveda y Rodero, 2003). Para confirmar la presencia de celo también fue utilizado un macho recelador. Cada vaca en celo fue pesada usando una romana (Morse & Co, EUA) y le fue insertado en la vena yugular externa, un catéter (Intracah 14 x 5½”) para la toma de muestras, la inyección de la droga o el diluyente. Los animales fueron divididos al azar en dos grupos. El grupo tratado (n = 6), después de una primera muestra de sangre, recibió ketamina (Sigma Chemical, EUA) diluida en solución salina (0,85%) a la dosis de 1 mg/kg. La inyección IV se hizo lentamente en un volumen de 20 mL. Después del tratamiento se tomaron muestras de sangre cada 5 min durante 2 h. Las mismas fueron colectadas en tubos Vacutainer® con EDTA y transportadas al laboratorio en una cava con hielo. En el grupo control (n = 6) se hizo mismo procedimiento, pero las vacas recibieron solamente igual volumen de solución salina al 0,85%. Las muestras de sangre fueron centrifugadas a 3000 rpm (Centrífuga Clay Adams, modelo 21152, EUA), durante 20 min. El plasma obtenido fue guardado por duplicado a –70°C en un freezer (Revco Scientific, EUA) hasta su procesamiento. Para la cuantificación de la LH las muestras fueron descongeladas a temperatura ambiente, minutos antes de su procesamiento. La LH fue determinada en muestras duplicadas de 100 µL utilizando un kit comercial (Diagnostics ACS 180, Chiron Corp., EUA) y un sistema automatizado de quimioluminiscencia (Ciba-Corning ACS 180 plus, GMI Inc., EUA). Este inmunoensayo se fundamenta en la determinación de una emisión de luz asociada con la disipación de energía de una sustancia electrónicamente excitada con partículas magnéticas del sustrato dioxetano. La sensibilidad del ensayo fue de 0,001 – 0,011 mUI/mL. También fueron determinados los coeficientes de variación intra-ensayos (6,38%) y el inter-ensayos (7,35%). Los valores fueron analizados usando la prueba de t de Student para la comparación de las medias, siendo consideradas significativas para P<0,05. RESULTADOS Los animales toleraron la inyección intravenosa de ketamina sin signos de somnolencia y dos de ellos mostraron signos oculares de inquietud, que fueron asociados con la acción anestésica disociativa de la ketamina. En la Figura 1 se presentan las concentraciones séricas de la LH después de la administración de ketamina y los valores de la LH en el grupo control que sólo recibió solución fisiológica. Al minuto cero, los promedios de LH fueron iguales en ambos grupos, pero a los 5 min se encontró una disminución de LH altamente significativa (P<0,01) en el grupo que recibió ketamina, el cual se mantuvo bajo hasta los 50 min. Después de transcurridos 55 minutos y hasta la finalización del experimento a los 120 minutos, no se encontró una diferencia significativa entre ambos grupos; de hecho, a partir del minuto 80, las concentraciones séricas de LH en las vacas tratada volvieron al valor inicial del experimento y las concentraciones de LH se igualaron al del grupo control (Figura 1). DISCUSIÓN La ketamina es un conocido antagonista selectivo, no competitivo y voltaje dependiente de los receptores NMDA (Reynolds y Millar, 1990). Bajo condiciones experimentales, en este ensayo, se presentan evidencias que confirman el papel que tiene el glutamato en la liberación de la LH, ya que la administración de ketamina disminuyó de forma altamente significativa su secreción durante 50 minutos. A pesar de que el glutamato, como neurotransmisor en la regulación neuroendocrina ha sido estudiado en muchas especies, en vacas lecheras, este ha sido el primer intento y nos permite alertar sobre el cuidado que debemos tener cuando usamos drogas que puedan alterar la actividad del glutamato durante el celo. Nuestros resultados coinciden con experimentos similares, en otras especies. Por ejemplo, en hembras Rhesus machaques durante la fase luteal del ciclo estrual, la administración de NMDA a dosis de 10 mg/kg, causó un rápido incremento de las concentraciones de la LH en plasma y esta estimulación fue inhibida de manera significativa por la administración de los antagonistas CNQX y MK801 (Urbanski et al., 1997). También en bovinos machos de 24 semanas de edad, la administración de MK-801, un antagonista no competitivo de los receptores NMDA, a dosis única de 0,1 mg/kg, abolió la secreción de la LH (Shahab et al., 1995). Recientemente, se ha determinado in vitro la presencia de receptores no-NMDA en células productoras de GnRH (Garyfallou et al., 2006). Asimismo, se han encontrado neuronas glutamatérgicas que contienen transportadores de glutamato (VGLUT1-3) con vesículas sinápticas que acumulan L-glutamato, en los núcleos hipotalámicos del control de la LH pre-ovulatoria, como el núcleo preóptico y el área hipotalámica anterior (Hrabovszky y Liposits, 2008). El efecto de la ketamina sobre la secreción de la LH probablemente es en el hipotálamo inhibiendo la GnRH, ya que la administración directa del glutamato en la hipófisis no altera la secreción de las gonadotropinas (Shainker y Ticero, 1980). Experimentos in vitro, usando células hipotalámicas de ratas incubadas con análogos del glutamato, produjeron una estimulación dosis-dependiente en la neuro-secreción de la GnRH, lo que confirma el sitio de acción central (Bourguignon et al.,1997). Pero este efecto varía con la condición del animal, ya que en ratas en lactación, el hipotálamo parece ser refractario ante la estimulación con los aminoácidos excitatorios y en consecuencia, no hay aumento de la GnRH después de la administración de NMDA. Esto podría deberse a que el amamantamiento induce la activación de péptidos opioides o de GABA que pueden suprimir a las neuronas hipotalámicas secretoras de GnRH (Pohl et al., 1989). Probablemente para que el NMDA tenga un efecto sobre la secreción de la LH se requiere que exista un ambiente hormonal adecuado, caracterizado por altas concentraciones de los estrógenos y bajas concentraciones de la progesterona (Pinilla et al., 1998). Los esteroides gonadales potencian el efecto estimulatorio de los aminoácidos excitatorios sobre la secreción de la LH en roedores (Carbone et al., 1992) y en ovejas (Estienne et al., 1990). Las acciones del glutamato y el estradiol sobre la secreción de las gonadotropinas son integradas en neuronas hipotalámicas donde se expresan ambos tipos de sus receptores (Diano et al., 1997). Dado estos antecedentes en el presente estudio se decidió tomar las muestras en el día del celo, ya que en ese momento, el folículo secreta grandes cantidades de estrógenos y los niveles de progesterona son bajos y se alcanza el valor máximo en el pico de la secreción de LH, que efectivamente fue suprimido por la acción de la ketamina. CONCLUSIÓN En el presente estudio, la inyección endovenosa de una dosis única de ketamina en vacas originó una disminución altamente significativa de la LH durante 50 minutos y abre la posibilidad de que situaciones similares puedan ocurrir cuando se usan otras sustancias que puedan afectar la neuro-secreción de la GnRH y, por ende, de la LH. La comprensión farmacológica del sistema reproductivo puede evitar problemas en la eficiencia reproductiva de nuestros rebaños. AGRADECIMIENTOS Expresamos nuestra gratitud al Dr. Mario Baptista, del Laboratorio Clínico Delta y al personal de Estación Experimental “Manuel Salvador Yépez”, en especial al M.V. Carlos Montezuma. Este trabajo fue subvencionado por el Consejo de Desarrollo Científico, Humanístico y Tecnológico (CDCHT) de la Universidad Centroccidental “Lisandro Alvarado” LITERATURA CITADA
The following images related to this document are available:Photo images[zt09030f1.jpg] |
| |||||||||

{kind=link}