 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
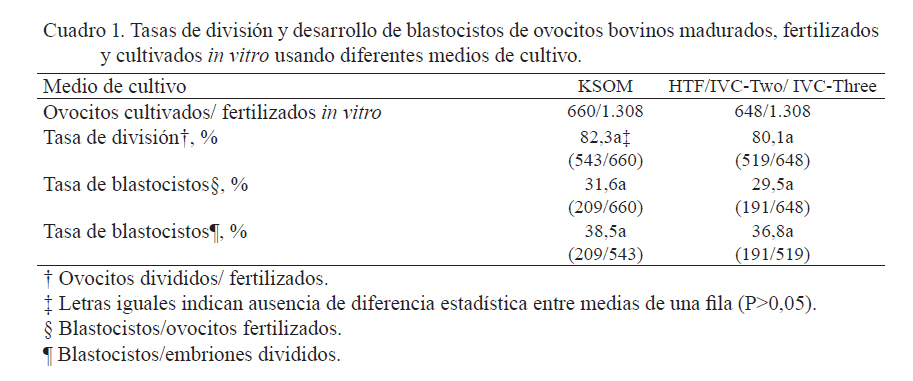
Zootecnia Tropical, Vol. 27, No. 3, 2009, pp. 277-284 Medio alternativo para la producción in vitro de embriones bovinos Alternative medium for in vitro production of bovine embryos Concepción Ahuja Aguirre1*, Felipe Montiel Palacios2, Ponciano Pérez Hernández1 y Jaime Gallegos Sánchez3 1Colegio de Postgraduados, Campus Veracruz. Carretera Federal Xalapa-Veracruz, C.P. 91700, Tepetates, Veracruz. México. *Correo electrónico: ahuja@colpos.mx Code Number: zt09031 RESUMEN Se evaluó la tasa de desarrollo de blastocistos bovinos producidos in vitro utilizando medio de cultivo convencional para embriones bovinos y medio de cultivo para producción in vitro (PIV) de embriones humanos. Se obtuvieron ovocitos de ovarios de vacas Cebú sacrificadas en matadero. Los folículos ováricos fueron aspirados y los complejos cumulus-ovocitos (CCO) con al menos dos capas de células del cumulus intactas fueron madurados durante 22 a 24 h. Posteriormente, los CCO fueron fertilizados con semen bovino descongelado y coincubados por 20 a 24 h. Los presuntos cigotos fueron cultivados por 48 h en medio KSOM o en medio HTF modificado. En los días 3 y 5 post-fertilización in vitro (FIV), los embriones fueron cambiados a gotas nuevas de KSOM o de IVC-Two/IVC-Three. Para ambos cultivos, en el día 3 post-FIV se determinó la tasa de división y entre los 7 a 9 días post-FIV, se evaluó el desarrollo embrionario a etapa de blastocisto. La tasa de maduración in vitro fue 89,7%. La tasa de división fue 82,3% para ovocitos cultivados en KSOM y de 80,1% para ovocitos cultivados en HTF (P>0,05). La tasa de desarrollo de blastocistos fue 31,6% para embriones cultivados en KSOM y 29,5% para embriones cultivados en HTF/IVC-Two/IVC-Three (P>0,05). En conclusión, las tasas de desarrollo in vitro de blastocistos bovinos fueron similares para los embriones cultivados en un medio convencional para PIV de embriones bovinos y para los embriones cultivados en medios utilizados para PIV de embriones humanos, por lo que ambos tipos de medios pueden utilizarse para la PIV de embriones bovinos con resultados aceptables. Palabras clave: Blastocisto, bovino, embrión, producción in vitro. ABSTRACT The objective of this study was to evaluate the blastocyst development rate for in vitro produced bovine embryos using either the conventional culture medium for bovine embryos or culture media used for the in vitro production (IVP) of human embryos. Oocytes were obtained from ovaries of slaughtered Zebu cows. Ovarian follicles were aspirated and the cumulus-oocytes complexes (COC) with at least two layers of intact cumulus cells were matured for 22 to 24 h. Following that, COC were fertilized with frozen-thawed bovine semen and coincubated for 20 to 24 h. The presumptive zygotes were cultured for 48 h in KSOM medium or modified HTF medium. On days 3 and 5, post-in vitro fertilization (IVF) embryos were placed into new drops of KSOM or IVC-Two/IVC-Three. For both culture types, the cleavage rate was determined on day 3 post-IVF and the embryo development rate to blastocyst stage was evaluated on days 7 to 9 post-IVF. In vitro maturation rate was 89.7%. Cleavage rate was 82.3% for oocytes cultured in KSOM and 80.1% for oocytes cultured in HTF (P>0.05). Blastocyst development rate was 31.6% for embryos cultured in KSOM and 29.5% for embryos cultured in HTF/IVC-Two/IVC-Three (P>0.05). In conclusion, in vitro blastocyst development rates were similar for both the embryos cultured in the conventional medium for IVP of bovine embryos and the embryos cultured in media used for the IVP of human embryos, thus, both types of media can be used for the IVP of bovine blastocysts with acceptable results. Keywords: Blastocyst, bovine, embryo, in vitro production. INTRODUCCIÓN La producción in vitro (PIV) de embriones bovinos ha adquirido importancia para ser usada como alternativa o en combinación con programas de ovulación múltiple y transferencia de embriones debido a sus ventajas y flexibilidad (Galli et al., 2001). El proceso de PIV de embriones bovinos se divide en cuatro etapas: la maduración de ovocitos, la capacitación espermática, la fertilización de ovocitos maduros y el cultivo de embriones. Estas etapas comprenden una serie de procesos fisiológicos, cada uno de los cuales condiciona el éxito o fracaso del proceso (Mucci et al., 2006). El parámetro más usado para medir la eficacia de la PIV de embriones es la tasa alcanzada de blastocistos (Kubisch et al., 2001). Aunque las tasas de maduración y fertilización in vitro de ovocitos bovinos son altas (aproximadamente 90 y 80%, respectivamente), la mayoría de los ovocitos fertilizados sólo se dividen hasta la etapa de 2 a 4 células y no todos son capaces de alcanzar la etapa de blastocisto (Lonergan et al., 1999; Rizos et al., 2002). La tasa de desarrollo de blastocistos in vitro oscila entre 30 y 40% (Marquant Le Guienne et al., 2001; Watanabe et al., 2001; Blondin et al., 2002; Lonergan et al., 2003a). Esto indica que el cultivo embrionario desde cigoto hasta blastocisto es crítico para la PIV de embriones, además que durante este período se define en gran medida, la calidad de los embriones obtenidos (Rizos et al., 2002; Galli et al., 2003; Lonergan et al., 2003b; Lonergan et al., 2004). Mientras el número de blastocistos PIV depende principalmente de la calidad de los ovocitos recuperados, la calidad de los blastocistos está determinada por el ambiente de cultivo y fertilización de los ovocitos (Calado et al., 2001; Holm et al., 2002; Rizos et al., 2002). Durante el cultivo embrionario in vitro ocurren cuatro eventos importantes en lo que se refiere al desarrollo desde la etapa de cigoto hasta la formación del blastocisto: la primera división embrionaria, cuyo momento de presentación es crítico para el subsecuente desarrollo del embrión (Lonergan et al., 1999), la activación del genoma embriónico en la etapa de 8 a 16 células (Memili y First, 2000), la compactación de la mórula en el día cinco (Boni et al., 1999) y la formación del blastocisto a los días seis ó siete (Watson, 1992). Por lo tanto, las condiciones inadecuadas del ambiente de cultivo que pudieran afectar alguno o todos estos eventos podrían tener un efecto deletéreo sobre la calidad del embrión (Lonergan et al., 2003b). Entre los medios más usados para el cultivo in vitro de embriones bovinos cabe mencionar: el medio Charles Rosenkrans (CR1) (Rosenkrans y First, 1994), el fluido oviductal sintético (SOF) (Gardner et al., 1994; Holm et al., 1999) y el medio optimizado simple de potasio (KSOM) (Liu y Foote, 1995). En México, en ocasiones resulta complicado conseguir medios para la PIV de embriones bovinos porque deben ser importados directamente de los laboratorios que los producen. Esto resulta en largos períodos de espera e inadecuado almacenamiento de los medios durante el transporte, lo que provoca que los medios no sean aptos para ser usados en el laboratorio, elevando el costo de los embriones producidos. Sin embargo, dado el auge que ha tenido en el país la reproducción asistida en humanos, generalmente resulta más fácil y menos costoso conseguir medios utilizados para la PIV de embriones humanos, que aunque deben ser igualmente importados, se consiguen con compañías distribuidoras nacionales. Por lo tanto, el objetivo del presente estudio fue evaluar las tasas de desarrollo de blastocistos bovinos producidos in vitro utilizando un medio de cultivo convencional para PIV de embriones bovinos y medios de cultivo empleados para la PIV de embriones humanos. MATERIALES Y MÉTODOS Los embriones fueron producidos según el protocolo descrito por Rivera et al. (2004), con excepción de que se utilizaron algunos medios diferentes a los mencionados por dichos autores. La solución Tyrode Modificada (Modified Tyrode solution) usada para preparar las soluciones TALP (Tyrode albumin lactate pyruvate) IVF-TALP y Sp (Sperm)-TALP, de acuerdo con Rivera et al. (2004), así como el medio optimizado simple de potasio (potassium simplex optimized medium; KSOM) fueron adquiridos de Caisson Laboratories Inc. (North Logan, EUA). Los medios fluido oviductal humano (HTF) modificado con y sin HEPES y con antibióticos (penicilina-estreptomicina 100 y 50 µg/mL) fueron obtenidos de In Vitro S.A. (Ciudad de México, México). La hormona folículo estimulante (FSH; Folltropin®-V) fue obtenida de Bioniche Animal Health (Ontario, Canada). Los medios IVC-Two™ (InVitroCare®; bajo en glucosa, libre de fosfato, formulado con EDTA, taurina y glutatión y adicionado con glutamina estabilizada), IVC-Three™ (InVitroCare®; libre de fosfato, con elevados niveles de glucosa, suplementado con aminoácidos esenciales y no esenciales, formulado con EDTA, glutatión y glutamina estabilizada) y la albúmina sérica humana (HSA) fueron adquiridos de InVitroCare, Inc. (Frederick, EUA). Se extrajo sangre de un bovino castrado para la obtención de suero de novillo, el cual fue procesado para desnaturalizar las proteínas y hacerlo apto para su uso en cultivo embrionario. Los demás reactivos fueron adquiridos de Sigma Aldrich Química (Toluca, México). El proceso de maduración, fertilización y cultivo de los embriones se realizó en una incubadora a una temperatura de 38.5°C en atmósfera de 5% de CO2 en aire húmedo. Colección y maduración in vitro de ovocitos Los ovarios fueron obtenidos de vacas Cebú sacrificadas en matadero. Inmediatamente después de su colección, los ovarios fueron colocados en un recipiente térmico con 1 L de solución salina estéril (0.9% [p/v] de NaCl) a 35-36°C suplementada con 100 UI/mL de penicilina G sódica y 1 mg/mL de estreptomicina. Al finalizar la colección, los ovarios fueron enjuagados para eliminar la sangre y fueron transferidos a otro recipiente térmico con nueva solución salina con penicilina/estreptomicina (35-36°C), donde fueron conservados para ser transportados al laboratorio (2 a 3 h después de su colección). Al llegar al laboratorio, los ovarios fueron lavados en solución salina a 38,5°C. Los CCO de los folículos ováricos de 3 a 7 mm de diámetro fueron aspirados con una aguja hipodérmica estéril calibre 18 unida a una jeringa desechable de 10 mL. El fluido folicular conteniendo los CCO fue transferido a una placa de Petri para seleccionar, bajo el estereomicroscopio (Leica Zoom 2000), sólo los ovocitos con al menos dos capas de células del cumulus intactas y con citoplasma granulado homogéneo. Los CCO seleccionados fueron colocados en el primer compartimiento de una placa de Petri de cuatro compartimientos (X-Plate™, BD Falcon™) conteniendo medio HTF-Hepes modificado con antibióticos (buffer HEPES 21 mM, bicarbonato de sodio 4 mM, sin albúmina humana fracción V, penicilina-estreptomicina 100 UI/mL y 50 µg/mL). Los CCO fueron lavados dos veces transfiriéndolos entre los compartimientos restantes de la placa. Posteriormente, los CCO fueron transferidos a una placa de Petri conteniendo medio de maduración de ovocitos (MMO) contentivo de HTF modificado con antibióticos (bicarbonato de sodio 25 mM, sin albúmina humana fracción V, penicilina-estreptomicina 100 UI/mL y 50 µg/mL), suplementado con 10% (v/v) de suero de novillo (SN), 50 mg/mL de gentamicina, 20 mg/mL de FSH (Folltropin®-V), 1 mg/mL de estradiol, 0,2 mM de piruvato de sodio y 15 mg/mL de L-glutamina para un lavado final. Grupos de 10 a 20 CCO fueron transferidos a gotas de 50 µL de MMO cubiertas con aceite mineral, y fueron incubados durante 22 a 24 h. Cumplido este tiempo, la maduración de los ovocitos se determinó al evaluar el grado de expansión de las células del cumulus con un estereomicroscopio. Preparación del semen y fertilización in vitro (FIV) Después de la maduración, los CCO fueron removidos de las gotas de maduración y lavados una vez en HTF-Hepes modificado con antibióticos y en IVF-TALP. Grupos de 20 a 30 CCO fueron transferidos a cada pozo de placas de cuatro pozos conteniendo 600 µL de IVF-TALP por pozo. Se descongelaron dos pajuelas de semen de dos diferentes toros de raza Holstein y el semen fue lavado por centrifugación mediante un gradiente de Percoll® (45%/90% [v/v]) a 1000 g por 10 min. El pellet de espermatozoides fue recuperado del fondo del tubo, diluido con 10 mL de Sp-TALP y centrifugado a 200 x g por 5 min. El sobrenadante fue desechado y el pellet fue resuspendido en 100 µL de IVF-TALP para obtener una concentración espermática de 2 x 106 espermatozoides/mL. A cada pozo conteniendo CCO maduros e IVF-TALP se le agregaron 25 µL de suspensión de espermatozoides (para una concentración final de 1 x 106 espermatozoides/mL) y 25 µL de PHE (2 mM de penicilamina, 1 mM de hipotaurina y 250 mM de epinefrina en 0.9% [p/v] de NaCl). Los ovocitos y los espermatozoides fueron coincubados por 20 a 24 h. Cultivo embrionario Finalizado el período de incubación, los presuntos cigotos fueron removidos de los pozos de la placa de fertilización y denudados de las células del cumulus mediante aspiración repetida utilizando para ello, una pipeta con punta de 135 µm de diámetro. Los ovocitos libres de células del cumulus fueron lavados dos veces en HTF-Hepes modificado y fueron entonces asignados para el cultivo en KSOM (Rivera et al., 2004) o para el cultivo en medios utilizados para PIV de embriones humanos. El cultivo con KSOM se hizo de la siguiente manera: los presuntos cigotos se lavaron una vez en KSOM y fueron colocados en grupos de 20 a 30 en gotas de 50 µL de KSOM suplementado con 3 mg/mL de albúmina sérica bovina esencialmente libre de ácidos grasos (EFAF-BSA), 0,5 µg/mL de gentamicina y 25 µL de aminoácidos no esenciales, cubiertas por aceite mineral y fueron cultivados durante 48 h. El día tres después de la FIV, los embriones fueron transferidos a gotas nuevas de 50 µL de KSOM cubiertas por aceite mineral y fueron cultivados por 48 h. El día cinco después de la FIV, los embriones fueron transferidos a gotas nuevas de 50 µL de KSOM suplementado con 5 µL de SN a cada gota de cultivo; las gotas fueron cubiertas con aceite mineral y se incubaron durante 48 a 96 h. El cultivo con medios utilizados para la PIV de embriones humanos se describe a continuación: los presuntos cigotos se lavaron una vez en HTF modificado y fueron colocados en grupos de 20 a 30 en gotas de 50 µL de HTF modificado cubiertas con aceite mineral y cultivados por 48 h. El día tres después de la FIV, los embriones fueron transferidos a gotas de 50 µL de IVC-Two suplementado con 10% de HSA, cubiertas con aceite mineral y cultivados por 48 h. El día cinco después de la FIV, los embriones fueron transferidos a gotas de 50 µL de IVC-Three suplementado con 12% de HSA, cubiertas con aceite mineral y cultivadas por 48 a 96 h. Para ambos medios de cultivo, el día tres después de la FIV se determinó la tasa de división de los embriones y a los 7 a 9 días después de la FIV se evaluó el desarrollo embrionario a etapa de blastocisto usando un microscopio invertido (Olympus CKX31). Análisis estadístico Las diferencias en las tasas de división y tasas de blastocistos para los medios de cultivo utilizados se analizaron usando la prueba de Chi cuadrado y la exacta de Fisher, disponibles en el paquete estadístico SAS (Cody y Smith, 1991). Se consideró una probabilidad de P<0,05 como estadísticamente significativa. RESULTADOS En el presente estudio, la tasa de maduración in vitro (porcentaje de ovocitos con cumulus expandido del total de ovocitos puestos a madurar) para ovocitos bovinos fue 89,7% (1.308 ovocitos madurados/1.458 ovocitos colectados). La tasa de división (porcentaje de embriones que alcanzaron el estadio de cuatro o más células del total de ovocitos fertilizados in vitro) fue 82,3% para ovocitos cultivados en KSOM y 80,1% para ovocitos cultivados en medio HTF modificado (P>0,05; Cuadro 1). La tasa de desarrollo de blastocistos (porcentaje de blastocistos obtenidos del total de ovocitos fertilizados in vitro) fue 31,6% para embriones cultivados en KSOM y 29,5% para embriones cultivados en HTF modificado / IVC-Two e IVC-Three (P>0,05; Cuadro 1). DISCUSIÓN Los ovocitos bovinos inmaduros permanecen en profase meiótica I dentro de los folículos ováricos, pero cuando son removidos de éstos y cultivados in vitro bajo condiciones adecuadas, reanudan la meiosis espontáneamente y aproximadamente 90% de ellos alcanzan la etapa de metafase II (Watson et al., 2000; Hashimoto et al., 2002; Mucci et al., 2006). La completa maduración del ovocito implica no sólo la adquisición de competencia meiótica, sino también la maduración citoplasmática (Motlik y Fulka, 1986). La maduración del ovocito depende en gran parte de la presencia de las células del cumulus que lo rodean, las cuales llevan a cabo funciones nutritivas y reguladoras (Sutovsky et al., 1993; Cetica et al., 1999). Algunos factores secretados por las células del cumulus, tales como glicosaminoglicanos y hormonas esteroideas, intervienen en la maduración citoplasmática (Dode y Graves, 2002). La expansión de las células del cumulus es necesaria para la fertilización y desarrollo embrionario temprano (Salustri et al., 1996; Eppig, 2001). La tasa de maduración in vitro de ovocitos bovinos obtenida en el presente estudio usando medio HTF modificado fue comparable a las tasas reportadas por diversos autores usando Medio de Cultivo Tisular 199 (TCM-199), las cuales variaron de 68 a 75% (De los Reyes et al., 1999), 82 a 90% (Iwata et al., 2004), 86% (Liu et al., 2008) y 71% (Li et al., 2009). La elección del medio de cultivo para la PIV de embriones bovinos depende de su eficiencia en la producción de blastocistos y de la competencia para el desarrollo que éstos muestren (Kuran et al., 2002). Estudios realizados por Barnett y Bavister (1996) y Krisher et al. (1999) demostraron que el medio de cultivo puede afectar el metabolismo de los embriones resultantes. En el presente estudio, las tasas de división y de desarrollo de blastocistos fueron similares para el cultivo en KSOM y para el cultivo en HTF/IVC-Two/IVC-Three. En este sentido, Fernández et al. (2007) reportaron una tasa de fertilización de 72%, tasas de división de 27 a 38%, una tasa de desarrollo de mórulas de 13% y de desarrollo de blastocistos de 2,6%, utilizando como medio de cultivo HTF. Por su parte, Krisher et al. (1999) obtuvieron tasa de división de 85% para ovocitos cultivados tanto en medio de cultivo para embriones de hámster 6 (HECM-6), como en medio SOF y tasa de 84% para ovocitos cultivados en medio G1/G2. Estos mismos autores obtuvieron tasa de desarrollo de blastocistos de 45% en HECM6, 38% en SOF y 30% en G1/G2, del total de ovocitos fertilizados. Utilizando como medio de cultivo el SOF, Boni et al. (1999) obtuvieron una tasa de división de 78% y una tasa de desarrollo de blastocistos de 28%. Kuran et al. (2002) reportaron tasas de división de 62 a 76% y de formación de blastocistos de 30 a 36% del total de embriones divididos. Stojkovic et al. (2002) indicaron una tasa de desarrollo de blastocistos de 9 a 17%; Palomares Naveda et al. (2004) obtuvieron tasas de división de 25 a 45% y de desarrollo a blastocistos de 11 a 35%; Lonergan et al. (2003b) reportaron tasa de división embrionaria de 81,4% y tasa de producción de blastocistos de 20,9%; Iwata et al. (2004) reportaron tasas de división de 60 a 76% y tasas de desarrollo de blastocistos de 24 a 47%; Dhali et al. (2009) obtuvieron una tasa de división de 71% y de desarrollo de blastocistos de 29% y Li et al. (2009) reportaron una tasa de división de 70% y de desarrollo de blastocistos de 26%. Por otro lado, utilizando como medio de cultivo el CR1aa (medio CR1 adicionado con aminoácidos esenciales y no esenciales), Urrego et al. (2008) obtuvieron una tasa de división de 73% y de desarrollo de mórulas y blastocistos de 20%. Sin embargo, de acuerdo con McEvoy et al. (2000), el alcanzar la etapa de blastocisto no garantiza que el embrión resulte apto para desarrollarse, por lo que es esencial, que los embriones que alcancen la etapa de blastocisto sean de la mejor calidad posible para asegurar óptimos porcentajes de gestación después de su transferencia (Lonergan et al., 2003b). Es sabido que la calidad de los blastocistos PIV es menor a la de aquéllos producidos in vivo, ya que en comparación con éstos, los embriones PIV muestran diferencias ultraestructurales (Crosier et al., 2001; Fair et al., 2001) que pueden resultar del sistema de cultivo específico usado (Thompson, 1997; Abe et al., 1999). Los efectos detrimentales del cultivo in vitro bajo condiciones subóptimas están mediados por modificaciones en la expresión genética del embrión. Estos efectos pueden expresarse como blastocistos incapaces de soportar la criopreservación o de mantener una gestación, así como la producción de crías de talla anormal, como ha sido observado en rumiantes (Young et al., 1998) y que se ha asociado directamente con el cultivo in vitro post-fertilización (McEvoy et al., 1998). Lonergan et al. (2003b) indicaron una sensibilidad temporal de los embriones bovinos al ambiente de cultivo post-fertilización, manifestada en términos de calidad de los blastocistos producidos, concluyendo que el período de cultivo después de la fertilización (desde el día 1 al día 7) fue el más crítico para determinar la calidad del blastocisto. En el presente estudio se pudo determinar que los medios de cultivo utilizados para PIV de embriones humanos fueron igualmente eficientes en la producción de blastocistos bovinos, que uno de los medios comúnmente utilizados para este fin. Esto representa una alternativa para la PIV de embriones bovinos en los casos en que se dificulta conseguir los medios que esta técnica requiere, ya que si se puede tener acceso a medios utilizados en reproducción asistida en humanos, éstos podrían emplearse para la PIV de embriones bovinos obteniendo una respuesta similar a la que se conseguiría con los medios comúnmente usados para esta especie. CONCLUSIONES Las tasas de desarrollo in vitro de blastocistos bovinos fueron similares para los embriones cultivados en un medio convencional para PIV de embriones bovinos y para los embriones cultivados en medios utilizados para PIV de embriones humanos. Por lo tanto, éstos últimos podrían utilizarse como alternativa para la PIV de embriones bovinos. LITERATURA CITADA
Copyright © 2009 - Zootecnia Tropical The following images related to this document are available:Photo images[zt09031t1.jpg] |
| |||||||||

{kind=link}