 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
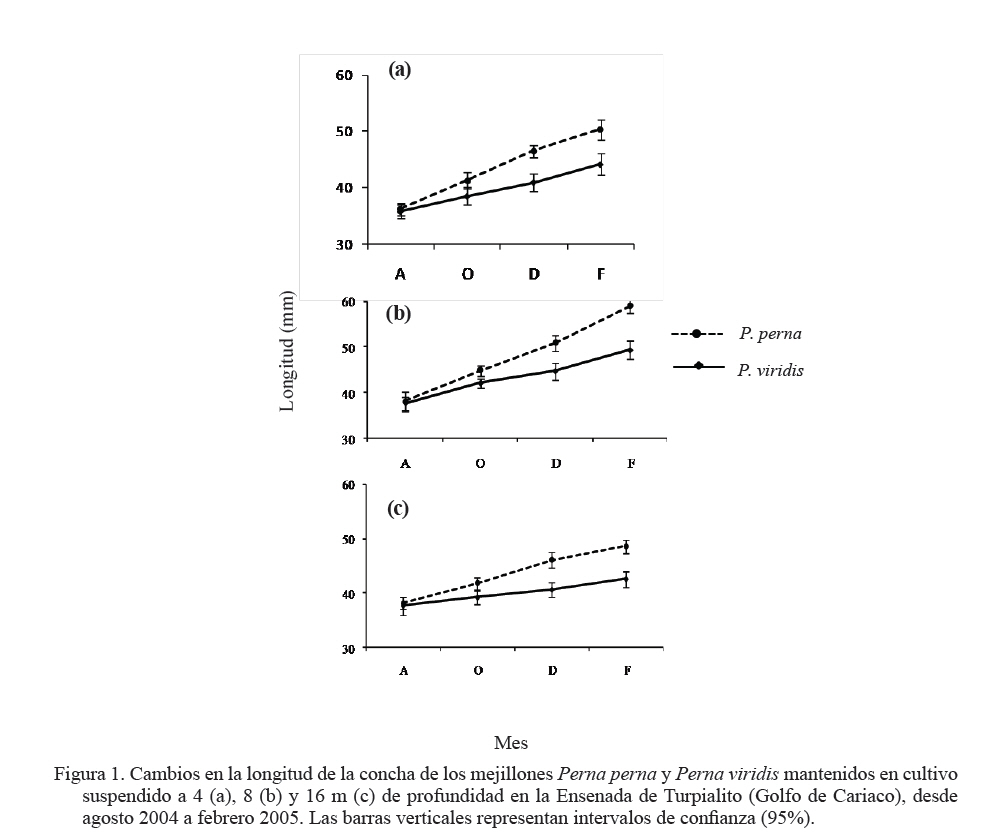
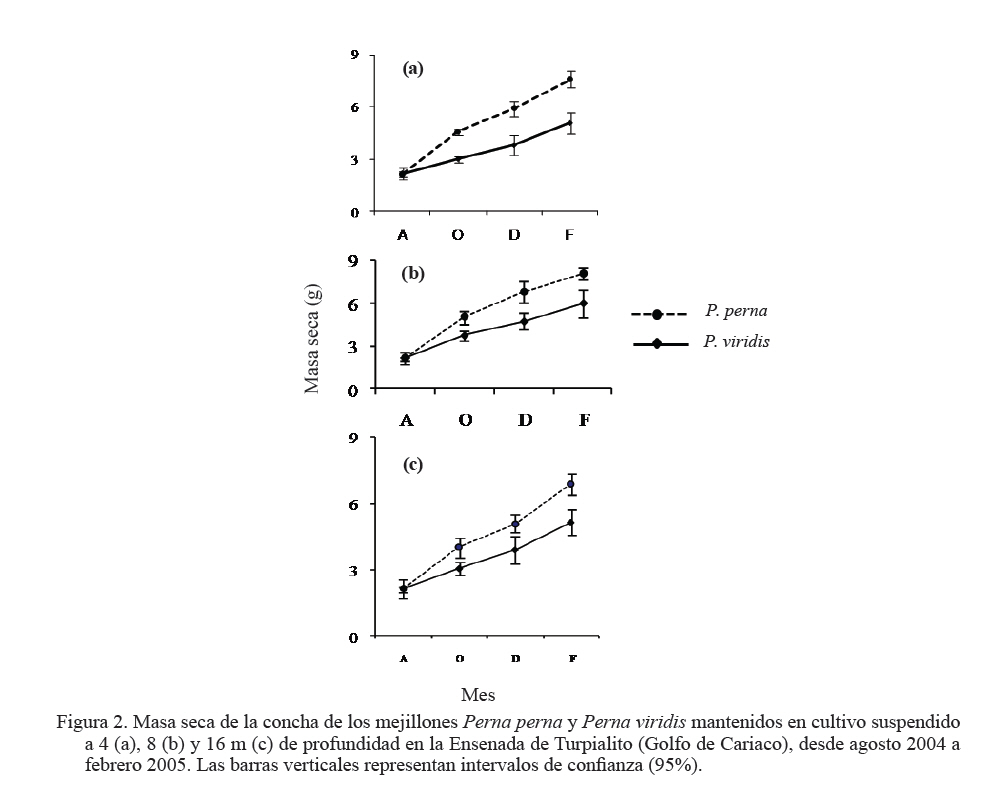
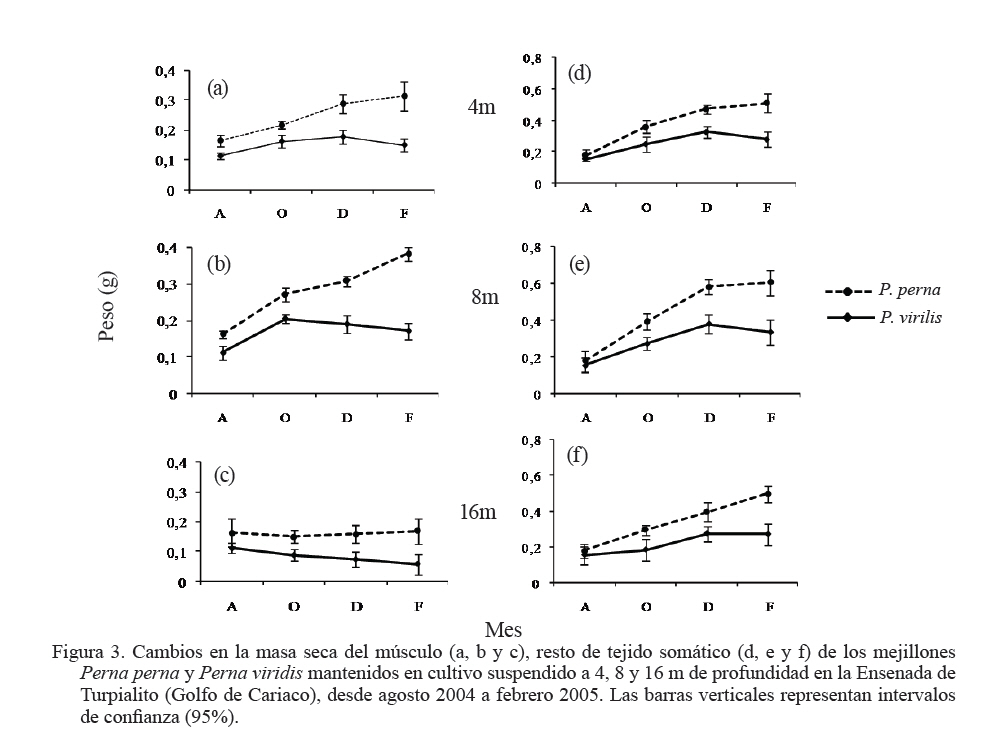
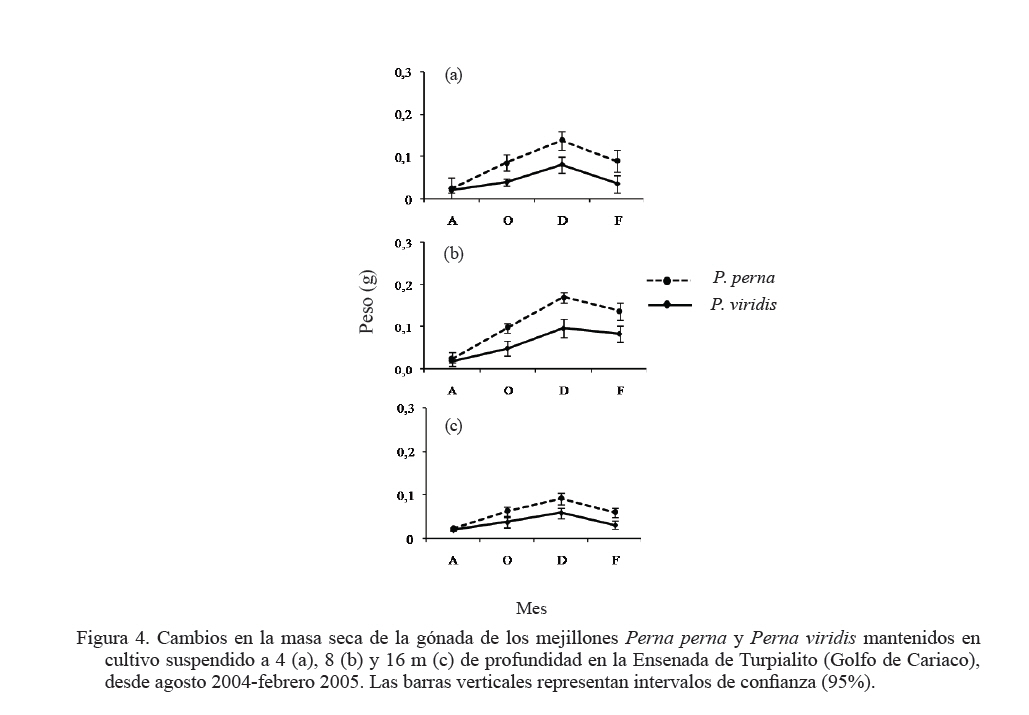
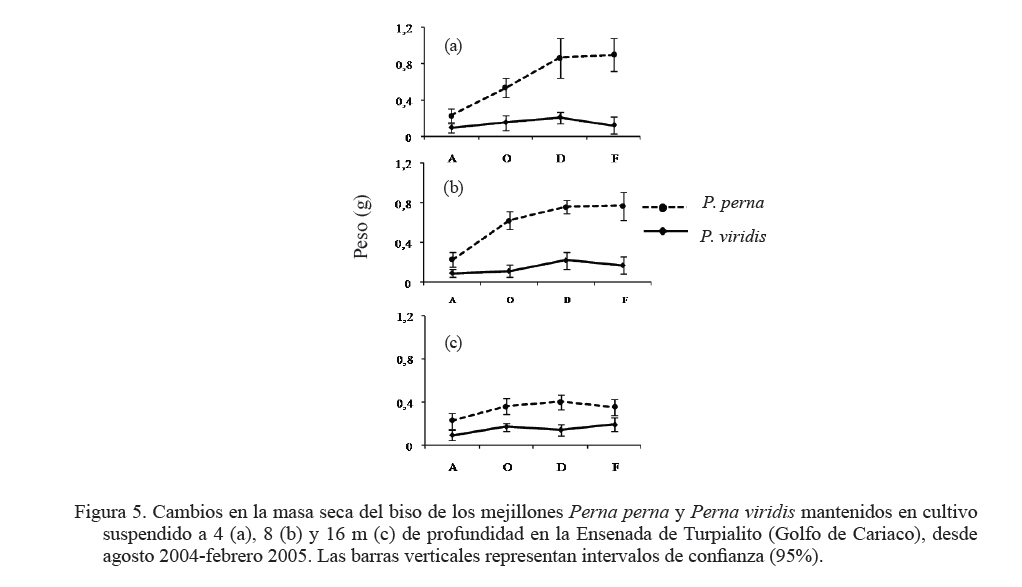
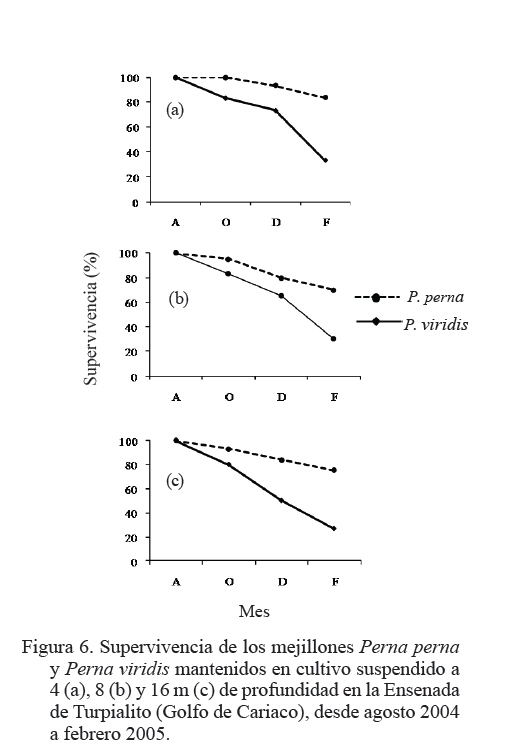
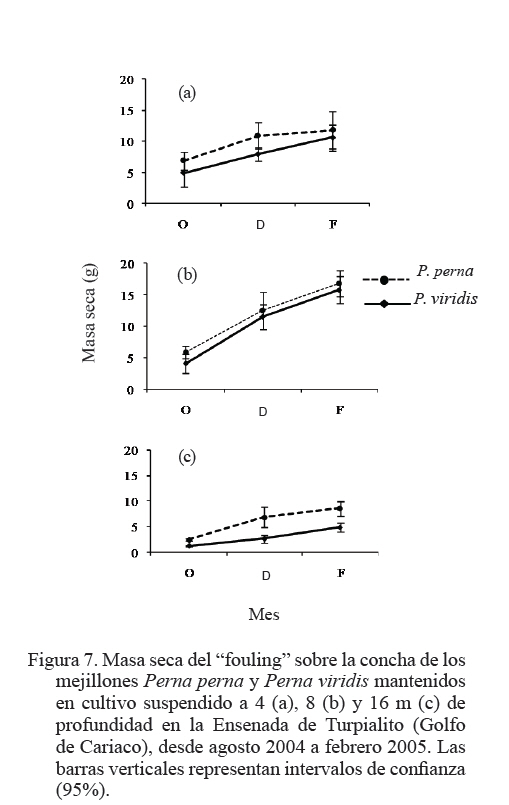
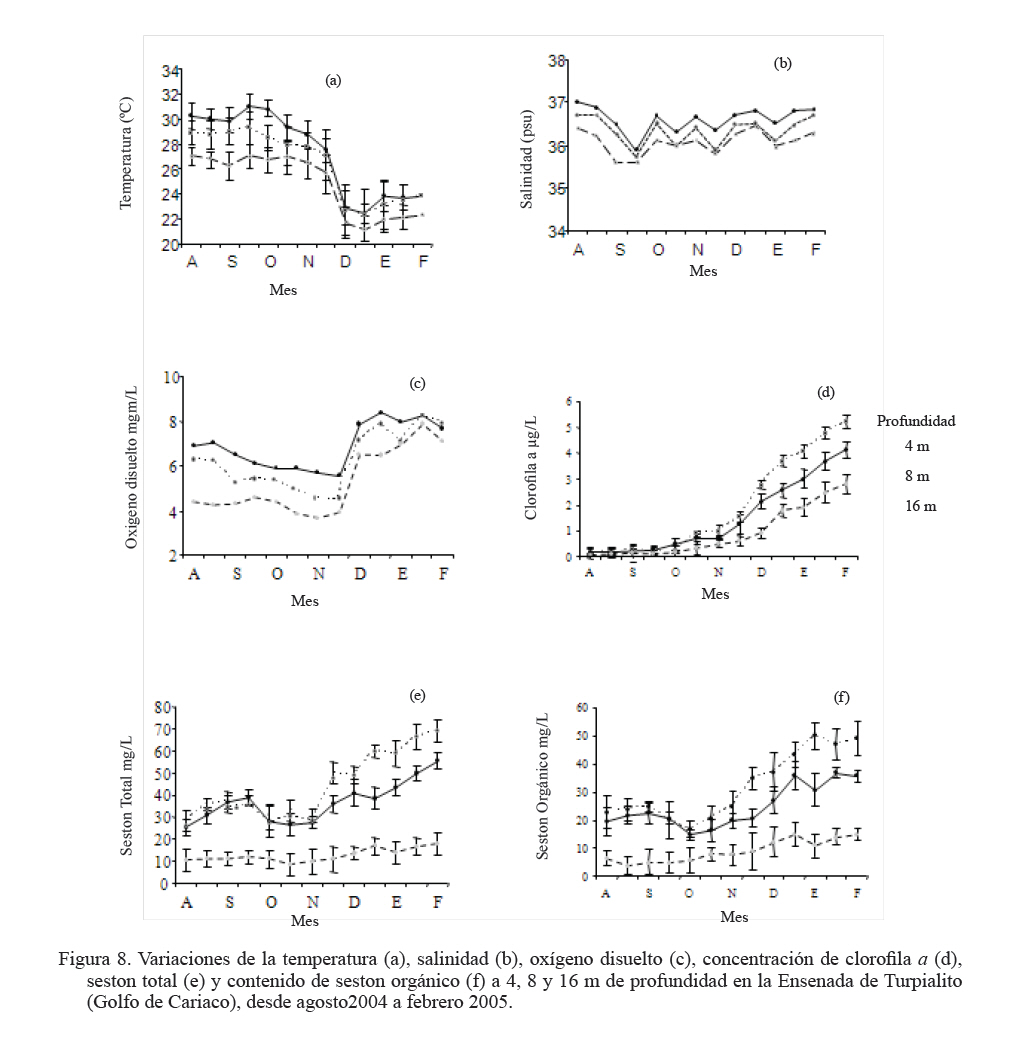
Zootecnia Tropical, Vol. 27, No. 3, 2009, pp. 315-328 Efecto de la profundidad sobre el crecimiento de los mejillones Perna perna y Perna viridis (Bivalvia:Mytilidae) en cultivo suspendido en el Golfo de Cariaco, Venezuela Depth effect on growth of the mussels Perna perna and Perna viridis (Bivalvia:Mytilidae) in suspended culture in the Cariaco Gulf, Sucre state, Venezuela Vanessa Acosta1*, Cesar Lodeiros2, Antulio Prieto1, María Glem1 y Yolimar Natera1 1Dept. Biología, Escuela de Ciencias. Universidad de Oriente. Cumana, Sucre. Venezuela. *Correo electrónico: vanessaacosta@yahoo.com Code Number: zt09035 RESUMEN Se evaluó el efecto de la profundidad (4, 8 y 16 m), sobre el crecimiento de los mejillones Perna perna y Perna viridis bajo condiciones de cultivo en el Golfo de Cariaco, Venezuela, durante un período de seis meses. Bimensualmente, se determinó la dimensión y masa seca de la concha, tejidos (somático y reproductivo) y el “fouling” sobre la concha en ambas especies. En cada profundidad se obtuvieron registros quincenales de temperatura, salinidad, oxígeno disuelto y disponibilidad de alimento (seston total, orgánico y biomasa fitoplanctónica). A los 8 m se alcanzaron los mayores incrementos en talla, masa seca de la concha, tejidos somático y reproductivo, obteniendo P. perna los mayores valores. El mayor crecimiento a esta profundidad estuvo asociado con el valor de seston orgánico (>20 mg/L). En ambas especies la masa del biso tendió a disminuir con el aumento de la profundidad, así como la biomasa del “fouling”. El menor crecimiento de P. viridis, conjuntamente con la baja supervivencia (<30%) en todas las profundidades experimentales sugiere la poca adecuación de esta especie para ajustarse a las condiciones ambientales presentes en el Golfo de Cariaco. Sin embargo, el continuo crecimiento de P. perna y la alta supervivencia (>70%) durante todo el periodo experimental demuestra que no fue afectada por los cambios ambientales que se produjeron en la columna de agua. De los factores ambientales, la disponibilidad de alimento fue el factor ambiental que determinó la diferencia de crecimiento entre las especies en las profundidades experimentales. Palabras clave: Cultivo, profundidad, Perna, Gofo de Cariaco, factores ambientales ABSTRACT The effect of depth (4, 8, and 16 m) on the growth of the mussels Perna perna and Perna viridis was evaluated under culture conditions in the Gulf of Cariaco, Venezuela, during a period of six months. Bimonthly, it was determined the dimension and dry mass of the shell, tissues (somatic and reproductive), and the “fouling” on the shell to both species. At each depth, biweekly registrations of temperature, salinity, dissolved oxygen, and food availability (total, organic seston, and fitoplanktonic biomass) were obtained. At 8 m depth, the highest increments were reached in size, dry mass of the shell, and somatic and reproductive tissues, obtaining P. perna the highest values. The higher growth at this depth was associated with the values of organic seston (>20 mg/L). In both species the mass of the repeat tended to decrease with the increment of the depth, as well as the biomass of the fouling. The lower growth of P. viridis, jointly with the low survival (<30%) in all the experimental depths, suggests little plasticity of this species to adjust to the environmental conditions of the Gulf of Cariaco. However, the continuous growth of P. perna and the high survival (>70%) during the experimental period demonstrates that it was not affected by the environmental changes that took place in the water column. Among the environmental factors, food availability was the most determining on the difference of growth between the species in the experimental depths. Keywords: Culture, depth, Perna, Gulf of Cariaco, environmental factors. INTRODUCCIÓN La optimización del crecimiento y los niveles de supervivencia son parámetros determinantes para el cultivo comercial de moluscos bivalvos. En este sentido, se han realizado estudios con la finalidad de evaluar y conocer el efecto de una serie de variables asociadas con los sistemas de cultivo, entre las que se encuentran: densidad (Parsons y Dadswell, 1992; Côte et al., 1993; Acosta et al., 2000), “fouling” (Claereboudt et al., 1994a; Lodeiros y Himmelman, 1996), acción de las olas (Widman y Rhodes, 1991; Freites et al., 1999) y la profundidad (MacDonald y Bourne, 1989; Emerson et al., 1994; Lodeiros et al., 1998; Kang et al., 2003; Ngo et al., 2006), siendo éste último uno de los parámetros más evaluados, debido a que los factores ambientales tales como la temperatura, salinidad, oxígeno y disponibilidad de alimento varían con el aumento de la profundidad causando efectos en el crecimiento, supervivencia y reproducción de algunos bivalvos marinos, bajo condiciones de cultivo. La influencia de la profundidad sobre el crecimiento de bivalvos ha sido ampliamente estudiada en zonas templadas y regiones boreales (Brcelj y Shumway, 1991; Thompson y MacDonald, 1991) en donde se ha encontrado un bajo crecimiento cerca de la superficie (MacDonald y Bourne, 1989; Román et al., 1999), mientras que otros estudios han reportado una disminución del crecimiento por debajo de una profundidad crítica donde las condiciones de temperatura, disponibilidad de alimento y turbidez a menudo exhiben un gradiente vertical subóptimo (Fréchette y Daigle, 2002). En el Golfo de Cariaco, al nororiente de Venezuela, existen variaciones de la temperatura (Okuda et al., 1978; Mandelli y Ferráz Reyes, 1982) y la biomasa fitoplanctónica a través de la columna de agua (Ferráz Reyes, 1989). Esta variabilidad ambiental ha inducido a realizar estudios en la zona a diferentes profundidades en pectínidos con potencialidades de cultivo en la zona, como Nodipecten nodosus y Euvola ziczac (Bonmatti, 1994; Freites et al., 1996; Lodeiros et al., 1998; Lodeiros y Himmelman, 2000) encontrándose efectos significativos de la profundidad sobre el crecimiento y la reproducción de dichos bivalvos. En este sentido, las diferencias ambientales que se generan a través de la columna de agua, como producto de la variabilidad ambiental que caracteriza las aguas del Golfo de Cariaco, pueden generar cambios en el crecimiento y comportamiento reproductivo de especies de mejillones como Perna perna y P. viridis, las cuales presentan un gran potencial para ser cultivadas a gran escala en dicha zona. En el presente trabajo, se evaluó el crecimiento y supervivencia de los mejillones P. perna y P. viridis, bajo tres profundidades experimentales y su relación con los factores ambientales, para establecer estrategias de cultivo con la finalidad de obtener una mayor producción por área de cultivo. MATERIALES Y MÉTODOS Los mejillones P. perna y P. viridis fueron recolectados en el banco natural existente en Guayacán, Península de Araya, estado Sucre, Venezuela (10º42’ N, 63º46’ O), los cuales fueron posteriormente trasladados a la localidad de Turpialito, Golfo de Cariaco (10º27’302” N, 64º01’52” O), para su cultivo en cuerdas a diferentes profundidades (4, 8 y 16 m) por un periodo de 6 meses (agosto 2004 a febrero 2005). Para llevar a cabo este estudio, se seleccionaron individuos juveniles con tallas homogéneas y significativamente iguales (ANOVA, P>0,05), para P. perna (38,3 ± 2,13 mm) y para P. viridis (37,7± 1,46 mm), respectivamente. La topografía del fondo de la Ensenada de Turpialito revela un perfil vertical de una considerable inclinación, llegando a unos 45 m de profundidad a 300 m de la orilla, aproximadamente. En dicha zona se instaló un “long line” de 50 m de longitud en donde se colocaron un total de 80 cuerdas (40 correspondiente a cada especie) de 1 m de largo con lotes de 40 mejillones cada una, por un período de 7 meses (agosto 2004 a febrero 2005) a diferentes profundidades (4, 8 y 16 m). Los mejillones fueron fijados a las cuerdas de caucho mediante una malla 100% nylon (tipo NE 200, La Vieira). Bimensualmente se extrajeron tres cuerdas del “long line” de cada especie de mejillón con la finalidad de evaluar el crecimiento (concha y tejido) y la sobrevivencia en cada una de las profundidades experimentales. A cada organismo se le determinó la longitud antero-posterior de la concha (altura), utilizando un vernier digital de 0,01 mm de apreciación, así como la biomasa seca (70°C/48 h) del músculo, lóbulos gonadales, glándula del biso y resto de tejidos (glándula digestiva, branquias, pie, manto, etc.), con una balanza analítica de 0,001 mm de precisión. De igual manera, para evaluar la masa seca del “fouling”, éste fue desprendido de las conchas y colocado en una estufa (70°C/24 h). Cada 15 días se colectaron muestras de agua por triplicado en cada profundidad con una botella Niskin de 2 L de donde se obtuvieron submuestras para la determinación de la concentración del oxígeno disuelto mediante el método de Winkler (Strickland y Parson, 1972). De la misma botella se tomaron muestras de agua, las cuales fueron previamente filtradas con una malla de 153 µm de tamaño de poro. Para estimar la biomasa fitoplanctónica mediante la concentración de clorofila a y el seston en sus componentes total, inorgánico y orgánico, las muestras fueron pasadas por filtros Whatman GF/F de 0,7 µm de diámetro de poro, utilizando un equipo de filtración al vacío Millipore. La clorofila a se determinó por el método espectrofotométrico, mientras que el análisis del seston se realizó mediante técnicas gravimétricas descritas en Strickland y Parsons (1972). Los registros de salinidad fueron obtenidos utilizando un refractómetro de 1‰ de apreciación, mientras que la temperatura fue determinada con un termómetro de inmersión. Para analizar los parámetros de crecimiento (altura de la concha, masa seca de la concha, músculo, gónadas y resto de tejidos), supervivencia y masa seca del “fouling” se aplicó un ANOVA de dos factores, empleando como factores el tiempo (meses) y las especies de mejillones. Cuando se detectaron diferencias significativas, se aplicó una prueba a posteriori de Duncan, empleándose un nivel probabilístico de 0,05. Para diferenciar las curvas de crecimiento entre ambas especies en el tiempo, se compararon sus pendientes aplicando un análisis de covarianza (Ancova), según Sokal y Rohlf (1981). RESULTADOS Talla En líneas generales se establecieron diferencias altamente significativas en el crecimiento en talla entre P. perna y P. viridis en las profundidades, durante todo el período experimental (P<0,001). Ambas especies presentaron un mayor incremento del crecimiento a los 8 m con respecto a las profundidades de 4 y 16 m, el cual siempre fue superior en P. perna. Esta tendencia se mantuvo desde el inicio del estudio hasta alcanzar una talla final de 58,9 ± 2,9 mm, con un incremento total de la longitud de la concha de 20,7 mm. Por el contrario, P. viridis alcanzó un crecimiento final de 49,4 ± 2,1 mm, correlativo con un bajo incremento de la concha (11,7 mm). Los organismos expuestos a la profundidad de 4 m mantuvieron un crecimiento significativamente igual (P>0,05) entre agosto y octubre 2004. Sin embargo, a partir de diciembre 2004, el crecimiento de P. perna fue progresivo hasta el final del estudio (50,3 ± 2,7 mm), mientras que en P. viridis la longitud de la concha mostró poca variabilidad en el crecimiento, alcanzando finalmente una longitud de 44,1 ± 1,6 mm. A ésta profundidad se obtuvo un incremento de la concha de 14,2 mm para P. perna y de 8,4 mm para P. viridis, respectivamente. A la profundidad de 16 m ambos mejillones presentaron la menor tasa de crecimiento, observándose el mismo comportamiento antes descrito, con una diferenciación del crecimiento partir de diciembre 2004, siendo significativamente mayor en P. perna (P<0,05) alcanzando un incremento total de la concha de 10,4 mm, con una talla final de 48,5 ± 2,2 mm. El incremento de la concha en P. viridis fue de 4,9 mm (42,6 ± 1,9 mm), respectivamente (Figuras 1a, b y c). Masa de la concha Al igual que en la longitud de la concha, ambas especies presentaron diferencias significativas, en todas las profundidades experimentales (P<0,05). A los 8 m ambas especies presentaron los mayores incrementos en la masa seca de la concha, siendo significativamente mayor en P. perna con respecto a P. viridis (P<0,05; Figuras 2a, b y c). Los mejillones colocados a 8 m alcanzaron una masa superior con respecto a las otras profundidades experimentales, obteniendo P. perna al final de la experiencia una masa de 8,1± 0,7 g y P. viridis de 6,0 ± 0,6 g, respectivamente. A 4 m, P. perna alcanzó una masa seca de la concha de 7,6 ± 0,6 g, mientras que la de P. viridis fue 5,1 ± 0,5 g. A 16 m, la masa seca de la concha fue menor a lo largo del estudio en ambas especies con respecto a las otras profundidades experimentales (P. perna 6,9 ± 0,7 g; P. viridis 5,1 ± 0,5 g, respectivamente). Músculo En este tejido se encontraron diferencias significativas en las profundidades experimentales entre las especies durante todo el período experimental (P<0,05). Las mayores masas de los tejidos también se obtuvieron a los 8 m, seguidos de 4 y 16 m de profundidad, respectivamente, observándose diferencias altamente significativas en el crecimiento en ambos mejillones y entre las diferentes profundidades durante toda la experiencia (P<0,05). En el músculo, a los 8 m entre agosto y octubre de 2004 se generó un crecimiento significativo en ambas especies; sin embargo, éste fue mayor en P. perna y se hizo progresivo hasta el final de la experiencia, alcanzando una masa de 0,3 ± 0,05 g. Contrariamente en P. viridis, desde octubre de 2005, se produjo un descenso en el peso del músculo, el cual se mantuvo hasta el final (0,1 ± 0,04 g). A los 4 m, la masa seca del músculo en P. perna presentó un crecimiento progresivo (0,3 ± 0,03 g), mientras que en P. viridis mostró un descenso en febrero 2005 (0,1 ± 0,04). A los 16 m, aunque no se obtuvieron diferencias entre ambas especies, P. perna presentó los mayores valores (Figuras 3a, b y c). Con respecto al resto de los tejidos somáticos, ambas especies presentaron el mismo comportamiento antes mencionado, con diferencias altamente significativas en las profundidades entre ambas especies durante toda la experiencia (P<0,05. Figuras 3d, e y f). Tejido reproductivo Con respecto a las gónadas, éstas tuvieron comportamientos diferentes entre P. perna y P. viridis en las diferentes profundidades (P<0,05). En líneas generales, P. perna presentó valores significativamente mayores a los alcanzados por P. viridis (P<0,05). A los 8 m ambas especies alcanzaron masas gonádicas superiores con respecto a las profundidades de 4 y 16 m. Para el mes de diciembre y a 8 m, se evidenció un incremento del tejido reproductivo en P. perna (0,1 ± 0,02 g), con respecto al de P. viridis (0,1 ± 0,02 g). Por el contrario, el crecimiento de este tejido a los 4 m fue mayor (P<0,05) en P. perna (0,1 ± 0,03 g) con respecto al alcanzado por P. viridis (0,04 ± 0,002 g). Para el mes de febrero 2005, en ambas especies se produjo una disminución de la masa de la gónada, evidenciándose un desove, que fue mucho más pronunciado a los 8 m y siendo superior en P. perna. A los 16 m el tejido reproductivo de P. perna mostró un incremento significativo (P<0,05) en el mes de diciembre (0,1 ± 0,02 g), para posteriormente presentar un descenso en febrero 2005 (0,01 ± 0,001 g). Por el contrario, en P. viridis no se observó un crecimiento significativo (P<0,05) del tejido reproductivo durante la experiencia (Figuras 4a, b y c). Masa del biso Entre P. perna y P. viridis se establecieron diferencias altamente significativas (P<0,0001) en la masa del biso en todas las profundidades durante el experimento. En ambas especies la masa del biso tendió a disminuir con el aumento de la profundidad. La mayor producción de biso la presentó P. perna siendo significativamente superior (P<0,05) a la de P. viridis en todas las profundidades experimentales y con tendencia a incrementarse con el tiempo (Figuras 5a, b y c). Supervivencia El valor de supervivencia en P. perna se mantuvo por encima de 70% en todas las profundidades durante todo el período experimental. Por el contrario, en P. viridis la supervivencia presentó una mayor variabilidad a través del tiempo y las profundidades experimentales, estableciéndose diferencias altamente significativas en dichas variables (P<0,05; Figuras 6a, b y c). “Fouling” La masa seca del “fouling” mostró diferencias significativas entre las especies y el período experimental en todas las profundidades (P>0,05). A partir de octubre 2004, se evidenció la incidencia de organismos incrustantes (fouling) en las conchas de los mejillones. La mayor cantidad de “fouling” sobre la concha de ambos mejillones se observó a los 4 m (P. perna 11,8 ±3,0 g; P. viridis 10,6 ± 2,1 g) y 8 m (P. perna 16,8 ± 2,3 g; P. viridis 15,1 ± 2,2 g), el cual mantuvo una tendencia al aumento desde diciembre 2004 hasta febrero 2005 (período de surgencia costera), haciéndose significativamente igual para ambas especies (P>0,05). Los ejemplares mantenidos a 16 m fueron los que presentaron menor masa de “fouling” sobre su concha; no obstante, a esta profundidad también se evidenció una tendencia al aumento de la masa haciéndose mayor en P. perna durante todo el período experimental, alcanzando al final una masa de 8,7 ±1,5 g en P. perna y de 4,9 ± 2,0 g, en P. viridis, respectivamente (Figuras 7a, b y c). La pendiente en la relación del crecimiento expresada en los cambios de la longitud de la concha, masa seca de la concha, músculo, resto de tejido, biso y “fouling” resultó ser siempre significativamente mayor en P. perna en todas las profundidades experimentales (P<0,001) entre las mismas. Factores ambientales La Figura 8a muestra los valores de la temperatura del agua en las diferentes profundidades experimentales. Entre agosto y septiembre de 2004 se pudo observar una variación de ±1°C dentro de cada profundidad experimental, mientras que las diferencias de temperatura durante el mismo período entre las profundidades fue de ±3°C, con una fluctuación entre 26 y 29°C. A mediados de noviembre 2004, se observó una caída brusca de la temperatura en todas las profundidades de aproximadamente 5°C, para alcanzar en diciembre de 2004, valores de 22°C, coincidentes con el inicio de la surgencia costera que anualmente acontece en el Golfo de Cariaco (Lodeiros y Himmelman, 2000). Entre enero y febrero 2005 la temperatura se mantuvo por encima de 23°C a los 4 y 8 m, mientras que a los 16 m estuvo entre 21 y 22°C. La salinidad mostró un comportamiento similar en todas las profundidades, con valores entre 35 y 36‰, durante todo el período experimental (Figura 8b). El oxígeno disuelto en el agua disminuyó con el aumento de la profundidad y mostró una tendencia a incrementarse con el tiempo (Figura 8c). La biomasa fitoplanctónica, estimada por clorofila a, fue baja entre agosto y octubre de 2004 (<1 µg/L) en todas las profundidades (Figura 8d). A partir de diciembre se observó que a 8 m los niveles de clorofila a comenzaron a incrementarse con respecto a las otras profundidades experimentales, comportamiento que se mantuvo hasta el final del estudio, con valores superiores a los 4 µg/L. Para el mismo período, a 4 m se mantuvo por encima de 3 µg/L, en contraste con la profundidad de 16 m en donde los niveles de clorofila a se mantuvieron en 2 µg/L. Las concentraciones del seston total mostraron tendencias diferentes en las profundidades experimentales (Figura 8e). Entre agosto y octubre 2004, a 4 y 8 m se obtuvieron valores superiores a los 30 mg/L, mientras que para el mismo período a 16 m, los niveles se mantuvieron en 10 mg/L hasta el final de la experiencia. El seston orgánico mostró un comportamiento similar al del seston total. En agosto y octubre a 4 y 8 m se obtuvieron valores de 20 mg/L, los cuales mostraron posteriormente una tendencia al aumento, siendo éste siempre mayor a 8 m, alcanzando valores superiores a los 30 mg/L a partir de noviembre. Sin embargo, a 16 m se observó baja variabilidad, ya que los niveles se mantuvieron alrededor de 15 mg/L, durante todo el período experimental (Figura 8f). DISCUSIÓN Las diferencias observadas en cuanto al crecimiento en talla y biomasa de P. perna y P. viridis en las diferentes profundidades experimentales pueden atribuirse a la variabilidad de los factores ambientales que se produjeron en la columna de agua y a la capacidad fisiológica de cada una de las especies de ajustarse a las condiciones del medio. Los incrementos en talla y biomasa alcanzados a 4 y 8 m, desde el inicio de la experiencia, fueron correlativos con los niveles de seston orgánico registrados en la zona entre agosto y septiembre (>20 mg/L), el cual fue suficiente para soportar el crecimiento de ambos mejillones, tomando en consideración que los mismos se encontraban aún en una etapa juvenil, por lo que toda la energía obtenida estuvo canalizada hacia el crecimiento. Los valores de seston registrados en la zona coincidieron con los aportes nutritivos relativos al período de lluvia que se produce entre mayo y septiembre. Cranford y Grand (1990) señalan que aunque el fitoplancton constituye un importante recurso alimenticio para los bivalvos, el seston orgánico es una fuente de energía alternativa para estos en períodos cuando los niveles de fitoplancton son escasos. No obstante, ambos mejillones alcanzaron los mayores incrementos de la concha a 8 m, con una mayor producción de tejido somático y reproductivo. En el caso de P. perna, dicho comportamiento se mantuvo durante todo el período experimental, mientras que en P. viridis al final de la experiencia se observaron descensos significativos tanto en el tejido somático como en el reproductivo. Estudios realizados en el Golfo de Cariaco señalan que los máximos niveles de biomasa fitoplanctónica se encuentran por encima de 10 m, fenómeno que está relacionado con el establecimiento de la termoclina que se produce entre 10 y 15 m, dentro de una profundidad de 30 m (Ferráz Reyes, 1989; Freites et al., 1996; Marín et al., 2004). Esta característica ambiental explicaría la mayor disponibilidad de alimento a los 8 m y su influencia sobre el crecimiento de ambas especies. En el Golfo de Cariaco existen variaciones en la temperatura de hasta 1,75°C a través de toda la columna de agua, entre la superficie y los estratos inferiores (Mandelli y Ferráz-Reyes, 1982), en donde dicha variabilidad se hace más pronunciada durante la estratificación de la columna de agua. En este estudio se obtuvieron diferencias de ±3°C entre 4 y 16 m, específicamente en agosto/octubre 2004 (período de estratificación), mientras que a 4 y 8 m la diferencia fue menor (±1,8°C). Los rangos de temperatura (28–29 y 29-30°C a los 8 y 4 m, respectivamente) que se registraron en los primeros meses del estudio (agosto-octubre 2004) fueron favorables para el crecimiento de ambos organismos, específicamente para P. viridis, que aunque mostró un crecimiento menor al de P. perna, mantuvo un incremento en su biomasa durante dicho período de tiempo. En este sentido, Arrieche et al. (2007) demostraron que P. viridis alcanza una alta eficiencia de asimilación del alimento a temperaturas constantes de 26 y 30°C, mientras que a temperaturas variables la tasa de filtración se ve afectada. Por lo tanto, la temperatura es un parámetro ambiental que afecta la capacidad de filtración de P. viridis, mostrando así un comportamiento característico de una especie tropical, con un patrón diferente al de especies subtropicales como P. perna, que está más adaptada metabólica y fisiológicamente a los cambios que se producen en el Golfo de Cariaco. Por lo tanto, la acción combinada de la temperatura y la disponibilidad de alimento y sus respectivas variaciones fue lo que marcó la diferencia en el crecimiento de ambas especies en las diferentes profundidades a través del tiempo. Aparte de las condiciones ambientales, las diferencias en el crecimiento en los mejillones a 4 y 8 m pudieron estar también condicionadas con un posible estrés fisiológico provocado por la exposición de los mejillones a los movimientos que se producen a través de la columna de agua, debido a la incidencia de las olas, específicamente en períodos de mayor prevalencia de vientos, relativos a la surgencia costera (diciembre a enero 2004/2005), donde los organismos a 4 m fueron los más afectados por estar más expuestos. La perturbación física causada por el movimiento de la columna de agua sobre los sistemas de cultivo ha sido demostrada en otras especies, particularmente en pectínidos como Patinopecten yessoensis (Ventilla, 1982), Placopecten magellanicus (Macdonald y Bourne, 1989) y en el Golfo de Cariaco para en Euvola ziczac (Freites et al., 1999), determinándose un efecto negativo de esta condición ambiental sobre el crecimiento y supervivencia de dichos bivalvos marinos. La mayor producción de biso obtenido por P. perna entre 4 y 8 m se produjo como una estrategia para fijarse más fuertemente a las cuerdas de cultivo, ya que con el transcurrir del tiempo el efecto del movimiento del sistema de cultivo, por efecto de las olas se hizo más fuerte, debido a la mayor incidencia de vientos, específicamente durante la surgencia costera. Además, la presencia de un biso fuerte es una de las adaptaciones morfológicas que le ha brindado a P. perna la capacidad y adaptabilidad de colonizar zonas intermareales rocosas (Mcquaid y Lindsay, 2000). La alta producción de biso garantiza un mayor refuerzo de los mejillones a las cuerdas de cultivo, lo cual evita su posterior desprendimiento. En tal sentido, el alto valor de supervivencia obtenido por P. perna (> 70%), durante todo el período experimental pudo estar asociado entre otros factores con la alta producción de biso. Un hecho que soporta lo antes señalado es la disminución progresiva de la supervivencia en P. viridis, la cual estuvo relacionada posiblemente, con pérdidas desprendimiento de los mejillones de las cuerdas de cultivo) de los mejillones, por la baja producción de biso. La profundidad de 16 m se caracterizó por presentar una baja disponibilidad de alimento, con respecto a las profundidades de 4 y 8 m, haciéndose desfavorable este ambiente para el crecimiento de ambas especies de mejillones, específicamente para P. viridis, donde el crecimiento de talla, biomasa, actividad reproductiva, producción de biso fueron afectados, así como la supervivencia, la cual disminuyó progresivamente hasta el final de la experiencia, obteniéndose un 20% de organismos vivos a esta profundidad. Estos resultados coinciden con los establecido por Fréchette y Daigle (2002) quienes señalan que el crecimiento disminuye a una profundidad crítica, en la cual la disponibilidad de alimento y la temperatura a menudo alcanzan un umbral para los organismos, donde no se produce crecimiento. Otros estudios realizados en el Golfo de Cariaco también han reportado una relación inversa entre el crecimiento y el aumento de la profundidad, relacionado principalmente con la disponibilidad de alimento (Bommattí, 1994; Freites et al., 1996; Lodeiros y Himmelman, 2000), ya que a través de la columna de agua los niveles de clorofila a pueden reflejar cambios en la composición y talla de las especies (Andersern y Naas, 1993). Lodeiros y Himmelman (2000) señalan que en el Golfo de Cariaco profundidades entre 21 y 30 m constituyen una alternativa en determinados momentos para el cultivo de pectínidos como Euvola ziczac, debido a que dichas profundidades pueden impedir la presencia de organismos dinoflagelados, causantes de marea roja. Estos “blooms” han originado mortalidades masivas de Nodipecten nodosus cultivados a bajas profundidades (8 m) en el Golfo de Cariaco (Lodeiros et al., 2000; Acosta et al., 2000). De acuerdo a lo antes señalado, profundidades superiores a 8 m podrían emplearse como una estrategia de cultivo en períodos críticos de “blooms” fitoplanctónicos en el Golfo de Cariaco. Esta alternativa se hace valedera tomando en consideración que uno de los principales factores que condujeron al declive (económico y productivo) del cultivo de P. perna en el Golfo de Cariaco fue la falta de estrategias de manejo que permitirían contrarrestar lo problemas producidos por la incidencia de biotoxínas. El efecto de la profundidad sobre la actividad reproductiva ha sido documentada en bivalvos marinos, coincidiendo en señalar que las diferencias en la producción de gónadas está asociada principalmente a las diferencias en los recursos alimenticios a través de la columna de agua y con las variaciones de temperatura (Rupp et al., 2004; Ngo et al., 2006). El máximo desarrollo de las gónadas en ambas especies se produjo en diciembre 2004 a las profundidades de 8 y 4 m, coincidiendo con la surgencia costera, donde se obtuvieron altos niveles de clorofila a, seston orgánico y bajas temperaturas (22-23°C). Posteriormente, se pudo evidenciar un decrecimiento gonádico, sugiriendo un desove en ambas especies (febrero 2005). La producción de gónadas se mantuvo sin variabilidad alguna en los ejemplares colocados a 16 m, lo que sugiere que las condiciones ambientales a esta profundidad no permitieron el desarrollo total de las gónadas debido posiblemente a la falta de alimento para cubrir la alta demanda energética que implica la gametogénesis, la cual fue sustentada posiblemente, por los bajos niveles de clorofila a observados conjuntamente con los altos niveles de temperatura. Las diferencias en cuanto a la producción de gónadas entre ambas especies estuvieron relacionadas con la capacidad de cada especie de regular su capacidad de alimentación y tolerancia a las variaciones de temperatura. El “fouling” es un factor ambiental que también puede influir negativamente en el crecimiento de los bivalvos (Lodeiros y Himmelman, 1996). En este estudio se descarta el efecto del “fouling” sobre el crecimiento de los mejillones en las diferentes profundidades experimentales, lo cual coincide con lo reportado por Acosta et al. (2009) quienes señalan que el “fouling” no afecta el crecimiento de P. perna y P. viridis bajo condiciones de cultivo suspendido, debido a la posición vertical en la que los mejillones se disponen en las cuerdas, lo cual impide que los organismos que componen el “fouling” tengan efecto directo sobre la apertura y cierre de las valvas. Este señalamiento coincide con lo reportado por otros autores para P. perna quienes han señalado que este parámetro no tiene efecto sobre el crecimiento de ésta especie (Monteiro y Silva, 1995; Metri et al., 2002). Las concentraciones de oxígeno y salinidad manifestaron cierta variación a través de las diferentes profundidades con intervalos de 4-8 mg/L y 36-37‰, respectivamente, los cuales se encuentran dentro los límites que no afectan fisiológicamente a dichos mejillones. En P. viridis y P. perna los valores óptimos de concentraciones de oxígeno disuelto fueron mayores a 5 mg/L y salinidad mayor a 28‰ (Segnini et al., 1998; Segnini, 2003), los cuales se obtuvieron durante el período experimental. Estudios fisiológicos sobre el efecto de la concentración del oxígeno y la salinidad bajo condiciones controladas y su relación con la temperatura son necesarios para verificar la hipótesis antes señalada. El menor crecimiento de P. viridis, conjuntamente con la baja supervivencia en todas las profundidades experimentales sugiere la poca plasticidad de esta especie para ajustarse a las condiciones ambientales que se producen el Golfo de Cariaco. Sin embargo, el continuo crecimiento de P. perna durante todo el período experimental demuestra que es poco afectada por los cambios ambientales que se producen, a través de la columna de agua. La alta supervivencia (>70%) mantenida por P. perna en todas las profundidades experimentales indica que es una excelente especie que muestra un rápido crecimiento en condiciones de cultivo suspendido. No obstante, se recomienda el cultivo de esta especie por encima de los 8 m y hasta 12 m de profundidad, lo que permitiría aprovechar mejor la profundidad del ambiente, con un mayor rendimiento de cultivo. De los factores ambientales analizados, la disponibilidad de alimento mostró ser el factor ambiental que determinó el crecimiento entre las especies. AGRADECIMIENTOS Este estudio fue financiado por el Consejo de Investigación de la Universidad de Oriente, a través del proyecto CI/5-1001-1155/03. Los autores agradecen la colaboración técnica del Lic. Paulino Núñez. LITERATURA CITADA
The following images related to this document are available:Photo images[zt09035f1.jpg] [zt09035f4.jpg] [zt09035f6.jpg] [zt09035f7.jpg] [zt09035f5.jpg] [zt09035f3.jpg] [zt09035f8.jpg] [zt09035f2.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}