
|
African Crop Science Journal
African Crop Science Society
ISSN: 1021-9730 EISSN: 2072-6589
Vol. 9, Num. 3, 2001, pp. 517-525
|
African Crop Science Journal, Vol. 9. No. 3, pp. 517-525
BIOLOGICAL CHARACTERISTICS OF TOMATO MILD MOTTLE VIRUS A
POTYVIRUS ISOLATED FROM TOMATO AND THORN APPLE IN ETHIOPIA
YAYNU HISKIAS1, D. - E. LESEMANN and H.J. VETTEN
Federal Biological Research Centre for Agriculture and Forestry, Institute
for Biochemistry and Plant Virology, Messeweg 11-12, 38104 Braunschweig, Germany
1Address for correspondence: Ethiopian Agricultural Research Organisation
(EARO), P. O. Box 2003, Addis Ababa, Ethiopia
Received 31 August, 2000
Accepted 3 February, 2001
Code Number: cs01071
ABSTRACT
Two isolates of the virus 246/94 and 277/94, acquired from thorn apple (Datura
stramonium) and tomato (Lycopersicon lycopersicum (L.) Karst. Ex.
Farw., syn. esculentum), respectively, were characterised biologically
and serologically and compared with a local isolate of Potato virus Y
(PVY) isolated from tomato and other potyviruses and isolates infecting vegetables.
Both isolates of TMMV infected only 16 of 28 plant species inoculated mechanically
and induced indistinguishable symptoms. The most susceptible hosts were D.
metel, D. stramonium and Nicotiana glutinosa L. However, these
isolates differed from the PVY isolate by infecting Datura spp. and Solanum
demissum L., while the PVY isolate infected Chenopodium quinoa Wild
and Capsicum annuum L. Isolate 277/94 was transmitted non-persistently
by the aphid, Myzus persicae Sulz. from diseased tomato to virus-free
D. stramonium, D. metel L., N. glutinosa
and tomato plants and from these back to virus-free test plants of each species.
Purified particles of isolate 277/94 contained a single protein species with
a molecular weight of 39 kDa. Furthermore, in double antibody sandwich enzyme-linked
immunosorbent assay (DAS-ELISA) antiserum to isolate 277/94 reacted strongly
with a Yemeni isolate of TMMV (Y90/7), but did not react with any other potyvirus.
This clearly shows that the Yemeni and Ethiopian isolates of TMMV are similar
serologically.
Key Words: Aphid transmission, host range, potyvirus, serology, tomato
mild mottle virus
RÉSUMÉ
Datura stramonium et Nicandra physalodes Gaertn en Ethiopie. Deux
formes du virus 246/94 et 277/94, ont ete obtenue respectivement partir du Datura
stramonium et de tomates (Lycopersicon lycopersicum (L.) Karst. Ex.
Farw., syn. esculentum) infectees. Leurs compositions biologique et sérologique
ont été identifiées et comparées avec: d'autres
potyvirus, un virus présent dans les pommes de terre locales Y (PVY)
et isolé à partir des tomates, et d'autres formes de virus
présents dans des légumes contaminés. Les deux formes du
virus TMMV ont contaminé 16 des 28 plants inoculés mécaniquement,
provoquant des symptômes similaires. Les plants où le virus s'est
cependant le mieux développé sont: le D. metel, le D.
stramonium et le Nicotiana glutinosa L. Cependant, ces deux formes
different du virus PVY dans la mesure où ils ont contaminé le
Datura spp. et le Solanum demissum L, alors que le PVY a contaminé
le Chepodium quinoa Wild et le Capsicum annuum L. Le virus 277/94
a été transmis de manière non persistante par l'
Aphid, Myzus persicae Sulz. Des tomates contaminées vers du D.
stramonium, D. metel L., N. glutinosa et
des plants de tomate saints, puis de ces plants infectés vers d'autres
plans tests saints de chaque espèce. Des particules purifiées
du virus (277/94) contiennent une seule espèce de protéine avec
une molécule pesont 39 kDa. De plus, une enzyme sandwich d'anti-corps
immunosorbent assay (DAS-ELISA) antiserum permettant d'isoler le 277/94,
a réagi fortement avec la forme Yéménite du virus TMMV
(Y90/7). Ceci démontre clairement que les formes Yéménite
et éthiopienne du virus TMMV sont très proches d'un point
de vue sérologique.
Mots Clés: Transmission des aphides, gamme des hôtes,
le virus de la tomate milde mottle
INTRODUCTION
About 30 plant viruses belonging to 11 different taxonomic groups infect tomato
in different countries of the world among which members of the Potyviridae
predominate (Jones et al., 1991). Among the potyviruses, Potato virus
Y (PVY) (Jones et al., 1991), Tobacco etch virus (TEV) (Harold, 1970;
Jones et al., 1991), Pepper veinal mottle virus (PVMV) (Ladipo
and Roberts, 1977; Jones et al., 1991) and Peru tomato virus (PTV)
(Fernandez-Northcote and Fulton, 1980; Jones et al., 1991) have been
reported from tomato and other vegetables, while Eggplant green mosaic virus
(Ladipo, 1976) and Tomato mild mottle virus (TMMV) infect only tomato
(Walkey, 1992; Walkey et al., 1994). Tomato mild mottle virus was described
from Yemen (Walkey, 1992), a country adjacent to Ethiopia and has now been
found to be the dominant and widespread pathogen of tomato in Ethiopia both
in single infections and mixed infections with PVY (Yaynu et al., 1999).
In addition to tomato (Lycopersicon esculentum) the virus was also
found infecting thorn apple (D. stramonium) and N. physalodes
weeds growing in association with tomato crops, in adjacent open fields, along
river banks and lake sides. Walkey et al. (1994) compared the serological
relationship of TMMV isolated from tomato in Yemen with only few other potyviruses
that infect tomato and pepper and observed no cross-reactions between TMMV
and the potyviruses.
The widespread occurrence of TMMV in tomato in Ethiopia as reported recently
(Yaynu et al., 1999) suggests that there may be many sources of infection
and an efficient transmission mechanism of the virus. Thus, it is necessary
to identify the sources of infection and the transmission mechanisms in the
affected areas and in greenhouse. This paper presents and discusses the host
range, aphid vector transmission and some biological and serological characteristics
of TMMV isolates from tomato (L. esculentum) and thorn apple (Datura
stramonium) in Ethiopia. Additionally, the Ethiopian TMMV isolates
are compared with a Yemeni isolate of the virus and other potyviruses from
vegetables in Ethiopia and other countries.
MATERIALS AND METHODS
Virus isolates and host range. Isolates of TMMV; 246/94 from D.
stramonium and 277/94 from tomato were collected in 1994 in the rainy
and dry seasons at Koka and Zwai in the Rift Valley of Ethiopia, respectively
(Yaynu et al., 1999). Isolate PVY-356/94 was collected from tomato
from Melkasa also in the Rift Valley. All TMMV isolates were maintained in
Nicotiana glutinosa and PVY-356/94 in N. tabacum var.
White Burley by sub-culturing every 30 days. Infected leaves were also stored
frozen at -200C to prevent possible contamination in the greenhouse.
Extracts from two-week-old systemically infected leaves were prepared as described
earlier (Yaynu et al., 1999). The resulting extracts were inoculated
each onto 4 carborundum-dusted test plants of the 16 plant species listed
in Table 1 and also to Gomphrena
globosa L. (Amaranthaceae), Chenopodium amaranticolor
Coste & Reynier, C. foetidum Schrad., C. foliosum
Aschers (Chenopodiaceae), Cucumis sativus L. (Cucurbitaceae),
Zea mays L. (Gramineae), Phaseolus vulgaris L., Vicia
faba L. (Fabaceae), N. hesperis L., N. miersii
Remy Nr. 33, and N. sylvestris Speg & Comes (Solanaceae)
under greenhouse conditions. All plants were inoculated at the 2-3 leaf-stage.
The experiment was repeated once and only species infected in both experiments
were included in Table 1. In addition
to visual observation, the presence of virus in the plants was confirmed by
testing both inoculated and non-inoculated leaves by ELISA (Clark and Adams,
1977).
Aphid transmission. Isolate 277/94 was transmitted by Myzus
persicae from infected tomato plants cv. Linda to virus-free test plants
of tomato cv. Linda, D. stramonium, D. metel,
N. glutinosa and from these back to virus-free test plants of
each species. Apterous M. persicae were fasted for 3-4 hr, allowed
an acquisition access period of 5-10 min on 277/94-infected plants and transferred
to virus-free test plants of each species using 30 aphids per plant. The experiment
was repeated once and results recorded. Aphids were killed after an inoculation
access period of two days. Equivalent acquisition accesses to virus-free plants
of all species were included as controls.
Virion length determination. For particle length measurements, virions
were trapped by immunosorbent electron microscopy (ISEM) from extracts of
277/94-infected plants. The lengths of 100 particles of isolate 277/94 were
measured at a magnification of 50,000x directly in a Zeiss EM906 electron
microscope using an on-line attached image analysing system (Digivion, SIS,
Muenster, Germany) with histogram production and normal length calculation.
Cytopathology of infected tissues. Cyto-pathological effects induced
by isolate 277/94 in N. tabacum cv. Xanthi-NC were studied as
described by Koenig and Lesemann (1985). Freshly harvested leaves were cut
into pieces, prefixed in 2.5% glutaraldehyde, fixed in 0.5% osmium tetroxide
and washed overnight in 1% uranyl acetate, pH 4.3. Leaf tissues were dehydrated
by sequential periods in increasing acetone concentrations for 90 min and
in a 1:1 (v/v) mixture of acetone and Epon-812 for 3 h. Leaf pieces were embedded
in Epon-812, polymerised at 600C for 48 h and ultrathin sections
were made using an ultramicrotome (LKB Ultratome III). The ultrathin sections
were viewed and photographed in a Zeiss EM 906 transmission electron microscope.
Virus purification. Isolate 277/94 was passed through D. stramonium
to exclude from PVY and propagated in N. glutinosa. Leaves showing
mosaic symptoms were harvested 10-12 days after inoculation and stored frozen
until use. Frozen leaves were purified as described previously (Siriwong et
al., 1995; Ravi et al., 1997; Yaynu and Vetten, 2000), without
the sugar density gradients. The purity and concentration of virus preparations
were examined by electron microscopy and yields of purified viruses
estimated spectrophoto-metrically (UV scan at 200 - 400 nm) as shown in Noordam
(1973).
Immunological and serological studies. Antiserum to isolate 277/94
was produced in a rabbit by weekly intramuscular injections of 200-400 mg
of purified antigen emulsified in an equal volume of Freund's incomplete
adjuvant for six weeks. Serological relationship of the isolate with other
viruses was studied in DAS-ELISA in homologous and heterologous combinations
using a total of 10 antisera as described by Clark and Adams (1977).
Coat protein size determination. Molecular weight (MW) of the coat
proteins (CP) of purified isolate 277/94 and other local potyvirus isolates
(374/94 and 430/94) from hot pepper were determined by sodium dodecyl sulphate
polyacrylamide gel electrophoresis (SDS-PAGE) using a 4% stacking gel on a
12% resolving gel and the buffer systems of Laemmli (1970). Gels were run
in a vertical electrophoresis apparatus (Mighty small II, Hoeffer Scientific
Instruments, San Francisco). Purified virus preparations were denatured by
boiling at 950C for 5 min. Amounts equivalent to 0.3, 0.25, 0.20
and 0.25μg of purified preparations of isolate 277/94, 374/94 and 430/94,
respectively, were loaded into the slots and electrophoresed first at 80V
until the bromphenol blue reached the resolving gel and continued at 120V
until they reached the bottom of the gel.
RESULTS
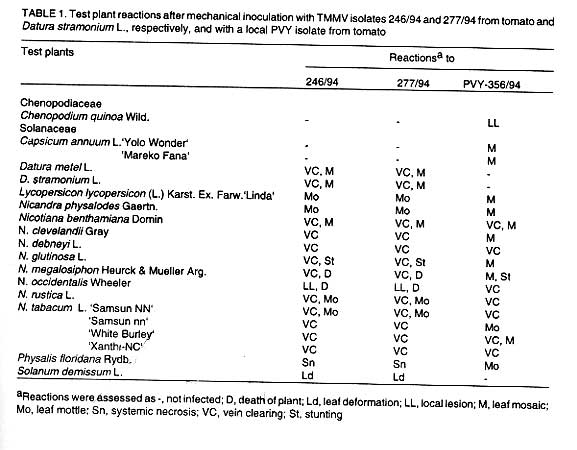
Experimental host range. The reactions of 16 test plant species to
mechanical inoculations with PVY-356/94, TMMV-246/94 and TMMV-277/94 are listed
in Table 1. The two TMMV isolates infected
16 solanaceous species on which symptoms were indistinguishable. All species
reacted with systemic symptoms except N. occidentalis, which developed
local lesions within 3 days of mechanical inoculation. The species highly susceptible
to both isolates were D. stramonium, D. metel, L.
lycopersicum cv. Linda, N. physalodes, N. benthamiana,
N. clevelandii, N. glutinosa and N. tabacum
vars. White Burley and Xanthi-NC of which the last two species were used for
virus multiplication and purification. These species developed various conspicuous
systemic symptoms 6-8 days after inoculation as compared to two weeks for those
inoculated by the PVY-356/94. Moreover, the majority of the species gave highly
specific DAS-ELISA readings with the TMMV-277/94 antiserum. Although many plant
species from different families were included in the host range study, the Ethiopian
isolates of TMMV only infected species in the Solanaceae, but not C.
annuum. Also N. miersii, N. sylvestris and N.
hesperis were further solanaceous species infected by the Ethiopian isolates
of TMMV. As shown in Table 1 the TMMV isolates
differed from the PVY isolate by infecting Datura spp. and S.
demissum, while the PVY - 356/94 infected C. quinoa and
hot pepper, which appeared to be immune to the TMMV isolates.
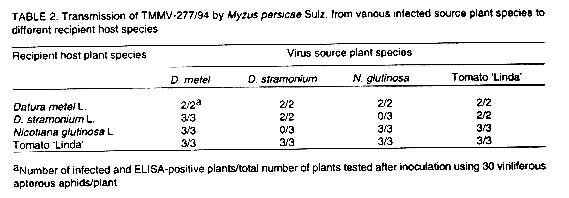
Aphid transmission. Isolate TMMV-277/94 was transmitted non-persistently
using all combi-nations of source and recipient host plant species, except from
N. glutinosa to D. stramonium and vice versa
( Table 2). Symptoms in N. glutinosa
and the two Datura spp. appeared 8-10 days after aphid inoculation,
whereas in tomato symptoms became visible after two weeks. The non-persistent
mode of transmission by M. persicae is characteristic of members
of the genus Potyvirus (Hollings and Brunt, 1981).
Particle morphology. Filamentous particles of c. 12 nm in diameter with
a modal length of 700 nm were observed consistently in leaf extracts infected
with isolate TMMV-277/94. This was unexpectedly short for a potyvirus, whose
normal length ranges from 680 to 900 nm (Hollings and Brunt, 1981). This, however,
is consistent with a value of 719 nm for isolate TMMV Y90/7 from Yemen (Walkey
et al., 1994).
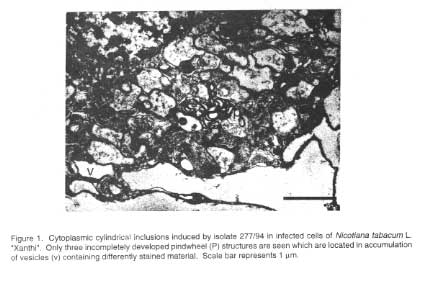
Cytopathological effects. As shown in Figure
1 cytoplasmic cylindrical inclusions (CI) were rare in TMMV-277/94-infected
tissue, which may be due to the extremely low particle concentration observed
in crude extracts. Moreover, the few CIs found in the 277/94-infected cells
were incompletely developed such that a clear assignment to one of the five
CI types could not be made. However, it seemed that CIs belonged to either type
II or type IV as described previously (Walkey et al., 1994).
Properties of purified virus preparation. Virus yields of only 2-4 mg
per kg of leaves were obtained for isolate TMMV-277/94, indicating a very low
concentration of this isolate in the source plant. Purified preparation of the
isolate still contained a considerable amount of host protein assumed to be
fraction I ( Fig. 2). The purity of the
purified preparation was also examined by spectrophotometry and SDS-PAGE, the
latter of which confirmed the presence of impurities in the virion prepation
as shown in Figure 2. The purified virus
had a UV absorption spectrum typical of the nucleoprotein of potyviruses with
a maximum at 260 nm, a minimum at about 246 and a slight shoulder at 290 nm.
The A260 nm/A280 nm ratio of the isolate was 1.06, which is lower than the range
typical for potyviruses (Noordam, 1973). This might have been due to the presence
of impurities.
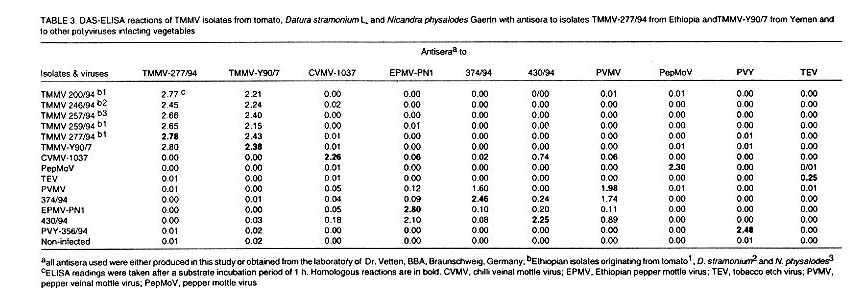
Serological relationship as determined by DAS-ELISA. The Ethiopian isolate
TMMV-277/94 was analysed in homologous and heterologous combinations by DAS-ELISA
using homologous antiserum and antisera to Yemeni isolate, Y90/97 as well as
eight antisera to other potyviruses infecting tomato and pepper locally and
other countries. The antisera to the Yemeni and Ethiopian isolates of TMMV reacted
not only in homologous and heterologous combinations, but also with four other
Ethiopian potyvirus isolates from tomato, D. stramonium and N.
physalodes, indicating that they are all serologically closely related
and can be regarded as TMMV isolates ( Table
3). None of the two TMMV antisera reacted with any of the eight other potyviruses
and their isolates, and no DAS-ELISA reactions were observed when the antisera
to different potyviruses and isolates were tested against the six TMMV isolates.
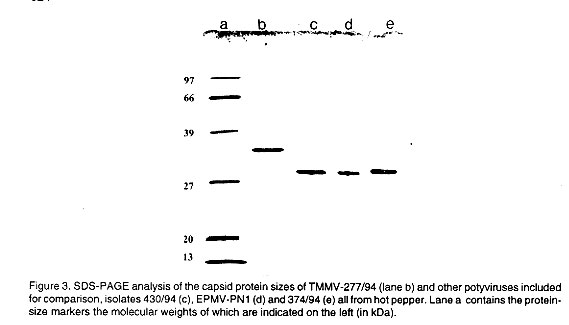
Capsid protein size. The SDS-PAGE analysis of purified preparation of
isolate 277/94 revealed that the isolate contained one major protein band with
M r of 39.0 ( Fig. 3 lane b).
However, the preparation also yielded, in addition to the major band, two minor
bands of c. 36 and c. 50 kDa. The former appeared to be a proteolytically degraded
form of the viral CP (Shukla et al., 1992), while the latter was regarded
as a protein co-sedimented with virions during purification ( Fig.
2). Isolate TMMV-277/94 had a CP size of c. 39 kDa which is in agreement
with a CP size of 42 kDa previously reported for a Yemeni isolate of the virus
(Walkey et al., 1994).
DISCUSSION
The incidence and severity of TMMV in tomato and associated solanaceous weeds
in many regions of Ethiopia were reported earlier (Yaynu et al., 1999),
but its infection route is not fully understood and must be determined in
order to devise appropriate control measures. Sixteen Solanaceous species
were identified as hosts of both TMMV isolates under greenhouse conditions.
Most of the hosts developed systemic infections, except N. occidentalis,
which produced local lesions to both TMMV isolates. This species can be used
as an assay host and for infectivity test with this virus. Most of the hosts
included in this experiment are wild or annual weed species that grow fast
and abundantly together with tomato in both rainfed and irrigated fields,
along river banks and lake sides throughout the year.
The natural vector(s) of TMMV is/are not yet known. However, under experimental
conditions the virus was transmitted from different infected plant species
to test plants by M. persicae in a non-persistent manner,
typical of potyviruses (Hollings and Brunt, 1981). This aphid species is known
to be polyphagous (Minks and Harrewijn, 1987) and is anholocyclic in the tropics
including Ethiopia, allowing rapid reproduction. Moreover, non-persistent
transmission by aphids provides fast virus spread from infected to healthy
plants in the field. The virus presumably over-winters on hosts such as D.
stramonium, and N. physalodes (Yaynu et al., 1999)
growing continuously along lake sides and river banks. To break the disease
cycle, over-wintering hosts growing together with tomato should be removed
so that vectors do not transmit the virus.
The formation of cytoplasmic cylindrical inclusions (CIs), also referred
to as pinwheels is the major criterion for the assignment of a new isolate
to the Potyviridae (Edwardson, 1974; Edwardson and Christie, 1996;
Shukla et al., 1992). The CIs induced by TMMV-277/94 were very rarely
found, which is probably due to extremely low particle concentrations as observed
in the crude extracts.
Different serological methods have been used to estimate the extent of cross-reactivity
among virus species of different groups (Clark and Adams, 1977; Koenig, 1978).
The DAS-ELISA method is highly specific and has been used to differentiate
between strains of the same virus, while indirect ELISA has been applied to
reveal distant relationships (Koenig, 1978; Koenig and Paul, 1982). An antiserum
was produced against isolate 277/94 of TMMV and its relationship with potyviruses
tested in homologous and heterologous combinations in DAS-ELISA. There was
no serological difference between the Ethiopian and the Yemeni isolate of
TMMV, while no cross reaction was observed between these and the other potyviruses.
This shows that TMMV is a serologically distinct virus as also shown by Walkey
et al. (1994).
Isolate 277/94 was conspicuous in having a CP size of c. 39 kDa, which is
largely in agreement with CP size of 42 kDa previously determined for the
Yemeni isolate (Walkey et al., 1994). Although SDS-PAGE revealed some
proteolytic degradation of TMMV CP, the apparent discrepancy between the size
determinations is probably due to different experimental conditions but not
due to proteolytic degradation of TMMV-277/94 CP. Moreover, the CP of the
Yemeni and Ethiopian isolates were indistinguishable in size when analysed
by electro-blot immunoassay (Vetten, personal observation). Another minor
band clearly observed above the major band could be due to impurities, co-sedimented
during purification.
REFERENCES
-
Clark, M.F. and Adams, N. 1977. Characteristics of the microtitre plate
method of enzyme-linked immunosorbent assay (ELISA) for the detection of
plant viruses. Journal of General Virology 34:475-483.
-
Edwardson, J.R. 1974. Some properties of the potato virus Y-group. Florida
Agricultural Experiment Station Monograph Series No. 4, 398 pp.
-
Edwardson, J.R. and Christie, R.G. 1996. Cylindrical inclusions. University
of Florida
Agricultural Experimental Station Bulletin 874, 79 pp.
-
Fernandez-Northcote, E.N. and Fulton, R. W. 1980. Detection and characterisation
of Peru tomato strains infecting pepper and tomato in Peru. Phytopathology
70:315-320.
-
Harold, F. 1970. Tobacco etch virus in Venezuela. Plant Disease Reporter
54:334-345.
-
Hollings, M. and Brunt, A.A. 1981. Potyviruses. In: Handbook of Plant
Virus Infections and Comparative Diagnosis.
-
Kurstak (Ed.). Elsevier, Amsterdam. pp. 731-807.
-
Jones, J.B., Jones, J.P., Stall, R.E. and Zitter, T.A. 1991. Compendium
of Tomato Diseases. American Phytopathological Society Press, St. Paul,
Mn. 132 pp.
-
Koenig, R. 1978. ELISA in the study of homologous and heterologous reactions
of plant viruses. Journal of General Virology 49:309-318.
-
Koenig, R. and Paul, H.L. 1982. Detection and differentiation of plant
viruses by various ELISA procedures. Acta Horticulturae 127:147-158.
-
Koenig, R. and Lesemann, D.-E. 1985. Plant viruses in German rivers and
lakes. Phytopathologische Zeitschrift 112:105-116.
-
Ladipo, J.L. 1976. A green mosaic disease of eggplant (Solanum melogena)
in Nigeria. Plant Disease Reporter 60:1068-1072.
-
Ladipo, J.L. and Roberts, I.M. 1977. Pepper veinal mottle virus associated
with a streak disease of tomato in Nigeria. Annals of Applied Biology
87:133-138
-
Laemmli, U.K. 1970. Cleavage of structural proteins during the assembly
of the head of bactriophages T4. Nature 227: 680-685.
-
Minks, A.K. and Harrewijn, P. 1987. Aphids: Their Biology, Natural
Enemies and Control. Vol. 2A. Elsevier Science Publishers. B.V. Amsterdam.
700 pp.
-
Noordam, D. 1973. Identification of Plant Viruses, Methods and Experiments.
Centre for Agricultural Publishing and Documentation, Wageningen, Netherlands.
207 pp.
-
Ravi, K.S., Joseph, J., Nakaraju, S., Krishna P. S., Reddy, H.R. and
Savithri, H.S. 1997. Characteristics of a pepper vein banding virus from
chilli pepper in India. Plant Disease 81: 673-676.
-
Shukla, D.D., Lauricella, R. and Ward, C.D. 1992. Serology of potyviruses.
Current problems and some solutions. In: Potyvirus Taxonomy. Barnett,
O.W. (Ed.). Archives of Virology 5: 57-69.
-
Siriwong, P., Kittipakorn, K. and Ikegami, M. 1995. Characterization
of chilli vein banding
mottle virus isolated from pepper in Thailand. Plant Pathology 44:718-727.
-
Walkey, D.G.A. 1992. Plant virus of Yemen and associated areas.
London, UK: Overseas Development Administration. 168 pp.
-
Walkey, D.G.A., Spence, N.J., Clay, C.M. and Miller, A. 1994. A Potyvirus
isolated from Solanaceous hosts. Plant Pathology 43:931-937.
-
Yaynu Hiskias, Lesemann, D.-E. and Vetten, H.J. 1999. Occurrence, distribution
and economic importance of potyviruses infecting hot pepper and tomato in
Ethiopia. Journal of Phytopathology 147:5-11.
-
Yaynu Hiskias and Vetten, H.J. 2000. Biological properties of potyviruses
from hot pepper (Capsicum annuum L.) in Ethiopia. Ethiopian Journal
of Pest Management 4:29-39.
© Copyright 2001, African Crop Science Society
The following images related to this document are available:
Photo images
[cs01071f3.jpg]
[cs01071f2.jpg]
[cs01071t3.jpg]
[cs01071f1.jpg]
[cs01071t1.jpg]
[cs01071t2.jpg]
|

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}