 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
African Crop Science Journal, Vol. 7. No. 4, pp. 333-339, 1999 Reactions of Musa genotypes to drought stress
J.A. Bananuka, P.R. Rubaihayo and M.M. Tenywa
Code Number: CS99023
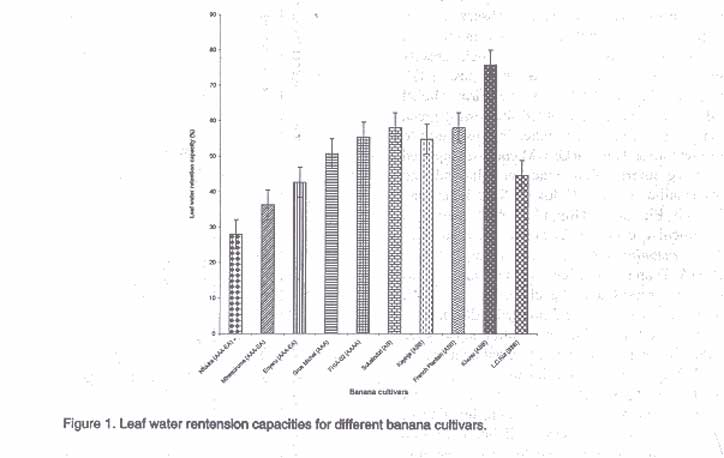
ABSTRACT Experiments were conducted at Makerere University Research Institute Kabanyolo (MUARIK), Uganda to identify the factors enabling the plant resistance and adaptability to drought stress in banana. Six banana cultivars (Nfuuka (AAA-EA); Sukalindizi (AB); French Plantain (AAB); Gros Michel (AAA); Lep Chang Kut (BBB) and FHIA-02 (AAAA) were subjected to 4.5 weeks of water stress during which soil moisture tension rose to over 400 mbar in potted experiments. Soil moisture tension of >400 (mbar) significantly (P<0.05) reduced growth, green leaf area, photosynthetic rate, stomatal conductance and sub-stomatal CO2 concentration. Cultivar Sukalindizi recorded significantly (P<0.05) lower transpiration rate and sub-stomatal CO2 but high assimilation rate indicating adaptability and resistance to drought stress, while cultivar Lep Chang Kut showed significantly (P<0.05) high transpiration rate and sub-stomatal CO2 concentration but low assimilation rate, leaf water retention capacity (44.5%) and stomatal conductance indicating its sensitivity to drought stress. The results from Lep Chang Kut were contrary to the concept that Musa balbisiana containing genomes have drought stress resistance. Gros Michel (AAA) and FHIA-02 (AAAA) exhibited stress resistance by retaining a high green leaf area (Y= -60.12x + 3261.5 and Y= -59.83x + 2558, respectively) and intermediate leaf water retention capacity (50.7 and 55.3%, respectively). Cultivar French Plantain (AAB) also exhibited intermediate moisture retention capacities (57.9%) indicating intermediate sensitivity to drought stress. The East African Highland banana Nfuuka had the lowest leaf water retention capacity (28.0%), indicating its sensitivity to drought stress.
Key Words: Banana, drought stress resistance, green leaf area, growth responses, stomatal conductance, transpiration rate
RÉSUMÉ
Des essais ont été menés à l’institut de recherche del’Université de Makerere, Kabanyoro (MUARIK), Uganda pour identifier de facteurs permettant l’adaptabilité et la résistance de plantes au stress due à la récheresse. Six cultivars de bananes (Nfuuka (AAA-EA); Sukalindizi (AB); French Plantain (AAB); Gros Michel (AAA); Lep Chang Kut (BBB) et FHIA-02 (AAAA) ont été soumis pendant 4 semaines et demie au stress d’eau jusqu’ à une tension d’humidité du sol de 400 mbar dans une experience en pots. La tension d’humîdité du sol supérieur à 400 mbar a réduit significativement (P<0.05) le croissance, la surface foliaire verte, le taux photosynthétique, la conductance de stomate et la concentration sous-stomatique du CO2. Le cultivar Sukalindizi a eu un taux le plus bas de transpiration, de dioxyde de carbone sub-stomatique, mais un taux d’assimilation élevé indiquant une adaptabilité et résistance au stress à la sécheresse, alors que le cultivar Lep Chang kut a eu un taux élevé de transpiration et de dioxyde de carbone sub-stomatique mais un taux bas d’assimilation, de capacité de retension en eau de la feuille (44.5%) et de conductance stomatique montrant une sensitivité au stress à la sécheresse. Les résultats de Lep Chang kut ont été contraires au concept que Musa balbisiana contenant de génomes possède une resistance au stress à la secheresse. Gros Michel et FHIA ont montré une résistance au stress en maintenant une surface foliairé verte élevée (Y = 60.12x + 3261.5 et Y = 59.83x + 2558 respectivement) et une capacité de retension en eau de la feuille intermediaire (50.7 et 55.3% respectivement). Les cultivars Kayinja et French Plantain ont eu des capacités de retension en eau foliaire de 54.7 et 57.9% respectivement montrant une sensitivité intermediaire au stress à la sécheress. La banane Nfuuka des hautes terres de l’Afrique de l’Est a eu la plus faible capacité de rétension en eau foliaire (28.0%), montrant sa sensitivité au stress à la sécheresse.
Mots Clés: Banane, résistance au stress à la sécheresse, surface foliaire verte, reponses à la croissance, conductance stomatique, taux de transpiration Introduction
Bananas (Musa spp.) rarely attain their full genetic potential for yield due to limitations imposed by water. The banana plants’ sensitivity to moisture stress is reflected in changes in reduced growth through reduced stomatal conductance and leaf size (Kallarackal et al., 1990; Turner, 1995), increased leaf senescence (Batalgia, 1980) ultimately limiting the plants’ photosynthesis. Robinson (1996) reported that soil drying in a healthy, previously well irrigated banana plantation to -12, -25, -53 and -70 Pa, induced a proportional reduction in photosynthetic rate amounting to 8, 18, 44 and 82 percent, respectively. Cayon et al. (1998) noted a 50% reduction in photosynthesis in cultivar "Dominico-Harton" under water stress compared to 30.61 µmol CO2 m-2 s-1 for unstressed plants. Leaf water retention capacity (LWRC %) was considered to be a better estimation of drought resistance than stomatal conductance (Clarke and McCaig, 1982). Tesha (1984) also observed LWRC to be more consistent than other methods. The main objective of the study reported in this paper was to determine the developmental and physiological processes that enable the banana plant to adapt to drought stress.
Materials and methods Tissue culture plantlets of six cultivars (Nfuuka (AAA-EA); Sukalindizi (AB); French Plantain (AAB); Gros Michel (AAA); Lep Chang Kut (BBB) and FHIA-02 (AAAA) were planted in buckets filled with soil with a layer of pebbles at the bottom. A tensiometer was inserted in each of the buckets to monitor soil moisture tension. The plants were watered for 4 weeks to allow for establishment after which water was withheld for 4.5 weeks. Soil moisture tension was measured with a pressure gauge 1000 (Agro-Technik, Bornheim, Germany). Leaf water retention capacity (LWRC %) was determined by the method described by Mukandawire et al. (1997). The third youngest leaf was removed from well watered plants and weighed within 10 minutes. The leaves were then hung on a wire to dry at room temperature for 48 hr, weighed and oven-dried at 800C for 48 hr. The amount of water retained during air drying (LWRC%) was obtained by subtraction and expressed as percent. Green leaf area was determined by measuring the length and width of all unfurled, fully expanded leaves before imposition of stress and repeated on weekly basis after imposition of water stress on newly unfurled leaves. Data were taken on length and width of the last emerged-fully expanded leaf, number of functional leaves (leaf with greater than 50% leaf area green), number of dry leaves (leaf with greater than 50% green leaf area senesced), and percentage of the leaf area still green for each leaf. Gauhl’s modification of Stover’s Sigatoka severity scoring method was adopted to score for the percentage of the leaf area that was still green. The percentage green leaf area at a particular time was given by the percentage of the leaf still green at a particular time multiplied by the leaf area. Leaf area was calculated using the formula reported by Jannoyer (1995): Leaf area = 0.83 (L x W). Measurement of photosynthesis rate (A), stomatal conductance (gs) and internal CO2 concentration (Ci) were taken with a portable photosynthesis system (Model CIRAS-1, Combined Infrared Gas Analyser, System Hitchin, Herts SG5 1RT UK), equipped with a PLC (B) leaf chamber calibrated to the manufacturer’s specifications. Leaf gas exchange measurements were taken on Mondays and Thursdays on the middle portion of the third youngest leaf between 1100 and 1300 hr on three plants. Plant height was measured from ground level to the point where the last two youngest leaf sheath separate, while pseudostem girth was measured 20 cm above soil level. Leaf emergence was taken after every two weeks and the total number of leaves produced and time to emit a single leaf determined. All data were subjected to analysis of variance using MstatC programme and means separated using LSD test.
Results and discussion The results of the effect of drought stress on plant growth presented in Table 1 show that drought stress of >400 (mbars) reduced pseudostem height and girth in all cultivars. Lep Chang Kut (BBB) recorded significantly (P<0.05) higher pseudostem height reduction, indicating its high sensitivity to drought stress, while Gros Michel (AAA) and Fhia-02 (AAAA) had the lowest reduction indicating resistance to drought stress. The girth reduction and number of leaves added did not significantly differ among cultivars. Turner (1995) noted that banana pseudostem girth and leaf emission are sensitive to water deficits. The results on green leaf area loss showed that the highest rate of green leaf area loss was observed in cultivars Lep Chang Kut (-455.21x) and Nfuuka (AAA-EA) (-309.13x) (Table 1) suggesting that they are senstive to drought stress. This observation for Lep Chang Kut is contrary to earlier findings that balbisiana genome cultivars have high capacity to resist drought (Stover and Simmonds, 1987). Cultivars FHIA-02 (-60.12x) and Gros Michel "Bogoya" (-59.83x) recorded the lowest green leaf area reduction rate, indicating resistance to drought stress. Cultivar Gros Michel, was reported to have intermediate reaction (Haddad and Manzanilla, 1995). Cultivars Sukalindizi and French Plantain "Gonja" (AAB) exhibited intermediate capacity. The better perfomance of Musa acuminata genome containing cultivars (FHIA-02 and Gros Michel) was not expected as this was reported to be characteristic of M. balbisiana (Stover and Simmonds, 1987). Table 1. Effect of drought stress on growth traits and green leaf area of different banana cultivars
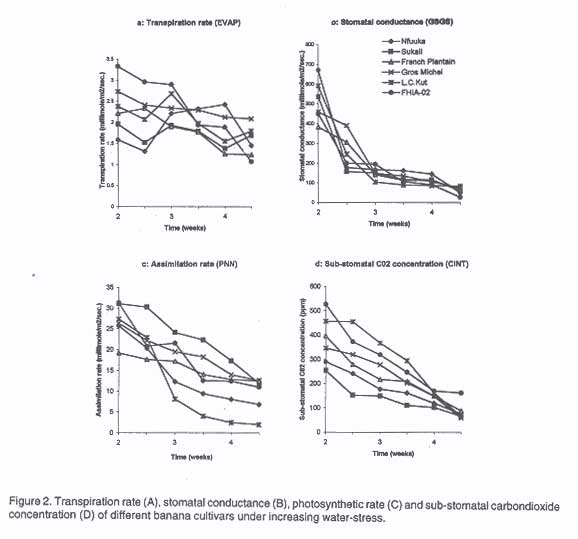
The results on leaf water retention capacity are shown in Figure 1. Cultivars exhibited significantly (P<0.05) different capacities to retain moisture after 48 hours of drying at room temperature. Cultivars FHIA-02, French Plantain, Sukalindizi, Kayinja (ABB) and Gros Michel exhibited significantly (P<0.05) higher leaf water retention capacity (51-58%) than other cultivars. The East African Highland banana Nfuuka had the lowest leaf water retention capacity (<43%) indicating their susceptibility to drought stress. The high water retention capacities of FHIA-02 and Gros Michel compared to Highland bananas explains their observed water stress resistance advantage. In wheat (Clarke and McCaig, 1982) and beans (Tesha, 1984), differences in leaf water retention capacity indicate cultivar differences to water loss and hence drought stress resistance. Balbisiana genome containing cultivar Lep Chang Kut had low leaf water retention capacity (44.5%) which explains its susceptibility to drought stress. The effect of drought stress of >400 (mbars) on transpiration rate, stomatal conductance, assimilation rate and sub-stomatal CO2 concentration are presented in Figure 2. Drought stress significantly (P<0.05) reduced transpiration rate in all cultivars (Fig. 2A). Cultivar FHIA-02 showed significantly (P<0.05) high reduction in transpiration rate (3.33 to 1.07 millimole/m2/sec.), while cultivar Nfuuka indicated an increase in transpiration rate (1.59 to 2.42 millimole/m2/sec. at 4th week) with increasing water stress which could explain its susceptibility to drought stress. The results on effect of drought stress on stomatal conductance presented in Figure 2B indicate a significant (P<0.05) reduction in stomatal conductance in all cultivars during the second and third week of stress. After the third week, cultivar Nfuuka maintained a higher stomatal conductance than the other cultivars until the fourth week when all cultivars showed a sudden fall. The maintenance of high stomatal conductance by Nfuuka explains its increase in transpiration rate. The results of drought stress of >400 (mbars) on assimilation rate are presented in Figure 2C. A reduction in assimilation rate of more than 50% was recorded in all cultivars, with exception of French Plantain (35.4%) by the end of 4.5 weeks. Cultivars Lep Chang Kut and Nfuuka recorded the highest reduction in assimilation rate (93.1 and 73.8 %), respectively. The high rate of leaf area loss by Lep Chang Kut and Nfuuka under drought stress explains their high reduction in assimilation rate. Cayon et al. (1998) noted a 50% reduction in photosynthesis in water stressed plants of cultivar "Dominico-Harton". Cultivars French Plantain, Gros Michel and Sukalindizi recorded and maintained significantly (P<0.05) low rates of reduction in assimilation rate, indicating their resistance to drought stress. de Ronde and van der Mescht (1997) noted that drought resistance is manifested by the ability of the crop to sustain a small reduction in physiological and/or metabolic activity under drought stress. Results on effect of drought stress on sub-stomatal CO2 concentration are shown in Figure 2D. Reduction in sub-stomatal CO2 with stress period was observed in all cultivars. Cultivar Sukalindizi exhibited low sub-stomatal CO2 concentration, while Lep Chang Kut had high sub-stomatal CO2 concentration, indicating low assimilation rates. The low sub-stomatal CO2 concentration in Sukalindizi was due to high assimilation rates as Farquhar et al. (1982) observed. Thus, increases in assimilation rate (A) relative to stomatal conductance (g) cause CO2 partial pressure (pi) to fall. Correlation analysis results indicate a relationships among the four characters (Table 2). Transpiration rate was positively correlated (P<0.01) to stomatal conductance and assimilation rate and negatively correlated to sub-stomatal CO2 concentration. Stomatal conductance was significantly (P<0.05) positively correlated to assimilation rate and sub-stomatal CO2 concentration, while assimilation rate was significantly (P<0.05) negatively correlated to sub-stomatal CO2 concentration. Similar observations were made by Robinson and Bower (1988) and Kallarackal et al. (1990), who reported a correlation between stomatal conductance, transpiration and photosynthesis in water stressed banana plants. Table 2. Correlation matrix for gaseous exchange parameters on bananas
The results suggest that Musa genotypes have different in-built mechanisms for resistance to drought stress. Cultivars which indicated small reductions in gaseous exchange and leaf area loss and high leaf water retention capacity and assimilation rates showed more resistance to drought stress.
Acknowledgement
The work reported in this paper was supported by Forum grant RF95007#75 from the Rockefeller Foundation. References
©1999, African Crop Science Society The following images related to this document are available:Photo images[cs99023a.jpg] [cs99023b.jpg] |
| |||||||||

{kind=link}
{kind=link}