 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Entomotropica, Vol. 16, No. 1, Abril/April, 2001, pp. 29-49 Neusa Hamada1, Marí Eugenia Grilleta 2 1Coordenação de Pesquisas em Entomologia, Instituto
Nacional de Pesquisas da Amazônia, Caixa Postal 478, 69011-970 Manaus,
AM, Brazil. E-mail: nhamada@inpa.gov.br. Code Number: em01003 ABSTRACT The present work provides distribution data and new identification keys to the larvae of 21 species of Simulium in the Gran Sabana region of southern Venezuela, including Canaima National Park. This work increases the black fly records in the area, providing the baseline needed for cytotaxonomic and ecological studies. Data were collected during three sampling periods (October 1996, February and October 1998) in Venezuela. Collections from the Pacaraima mountain region (Roraima, Brazil) on the Brazil-Venezuela border were also included. In the Gran Sabana the following species were collected: S. bipunctatum, S. cauchense, S. guianense s.l., S. goeldii, S. ignacioi, S. inaequale, S. incrustatum, S. iracouboense, S. kabanayense, S. lutzianum, S. maroniense, S. metallicum s.l., S. perflavum, S. quadrifidum, S. spinibranchium, S. suarezi and S. subpallidum. In Pacaraima the species were S. cauchense, S. covagarciai, S. lutzianum, S. maroniense, S. metallicum s.l., S. perflavum, S. rorotaense, S. rubrithorax and S. trombetense. Two new records are reported from southern Venezuela and Brazil: S. covagarciai and S. metallicum s.l. Simulium metallicum cytotype E in northern Venezuela, and S. guianense s.l. and S. incrustatum in southern Venezuela/northern Brazil are associated with onchocerciasis transmission. Future studies in the area will be necessary to determine cytotypes present and their biting behaviour. Additional key words: Aquatic insects, Neotropical region, simuliids, taxonomy. RESUMEN HAMADAN, GRILLET ME. 2001. Simúlidos (Diptera: Simuliidae) de la Gran Sabana (Venezuela) y la región Pacaraima (Brasil): datos de distribución y claves de identificación para larvas y pupas. Entomotropica Vol. 16(1):29-49. Este trabajo presenta nuevas claves de indentificación para larvas y pupas, y datos de distribución de 21 especies del género Simulium en la región de la Gran Sabana (Parque Nacional Canaima), sur de Venezuela. Incrementamos así la información previa existente y ofrecemos la base para estudios citotaxonómicos y ecológicos en el área. Las colecciones se hicieron durante tres períodos: octubre 1996, febrero y octubre 1998, y se incorporaron datos de la región fronteriza cercana de Brasil, Pacaraima (Roraima). Las especies colectadas en la Gran Sabana fueron: S. bipunctatum, S. cauchense, S. guianense s.l., S. goeldii, S. ignacioi, S. inaequale, S. incrustatum, S. iracouboense, S. kabanayense, S. lutzianum, S. maroniense, S. metallicum s.l., S. perflavum, S. quadrifidum, S. spinibranchium, S. suarezi y S. subpallidum. Las colectadas en Pacaraima fueron: S. cauchense, S. covagarciai, S. lutzianum, S. maroniense, S. metallicum s.l., S. perflavum, S. rorotaense, S. rubrithorax y S. trombetense. Presentamos dos nuevos registros para el sur del Orinoco (Venezuela) y Brasil:S. metallicum s.l. y S. covagarciai. Simulium metallicum citotipo E (norte de Venezuela), y S. guianense s.l. y S. incrustatum (Amazonas, Venezuela-Brasil) están asociados con la transmisión de oncocercosis humana. Trabajos futuros en el área de estudio deberían determinar los citotipos presentes y el comportamiento de picada de estas tres importantes especies. Palabras claves adicionales: Insectos acuáticos, región Neotropical, simúlidos, taxonomía. INTRODUCTIONThe family Simuliidae has a world-wide distribution far (Crosskey 1988), simuliid
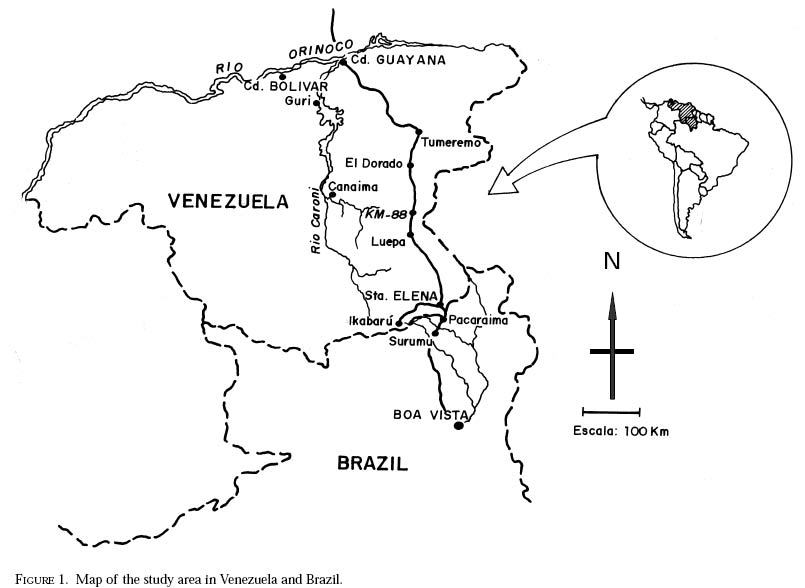
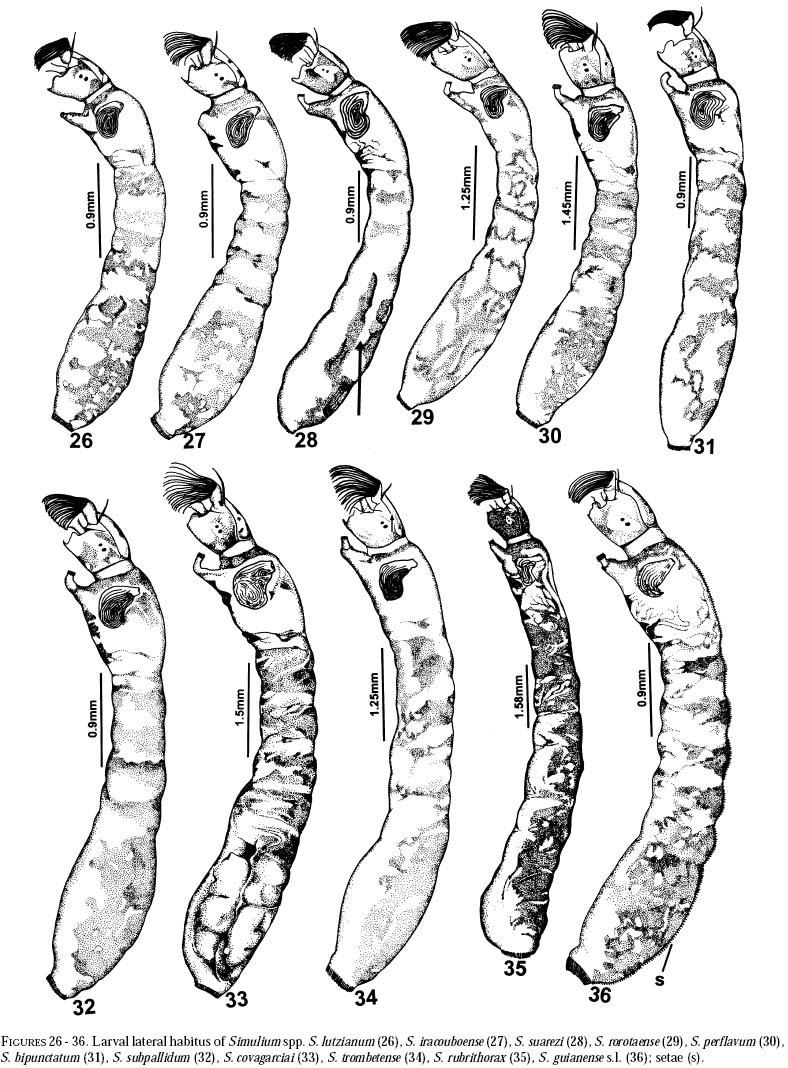
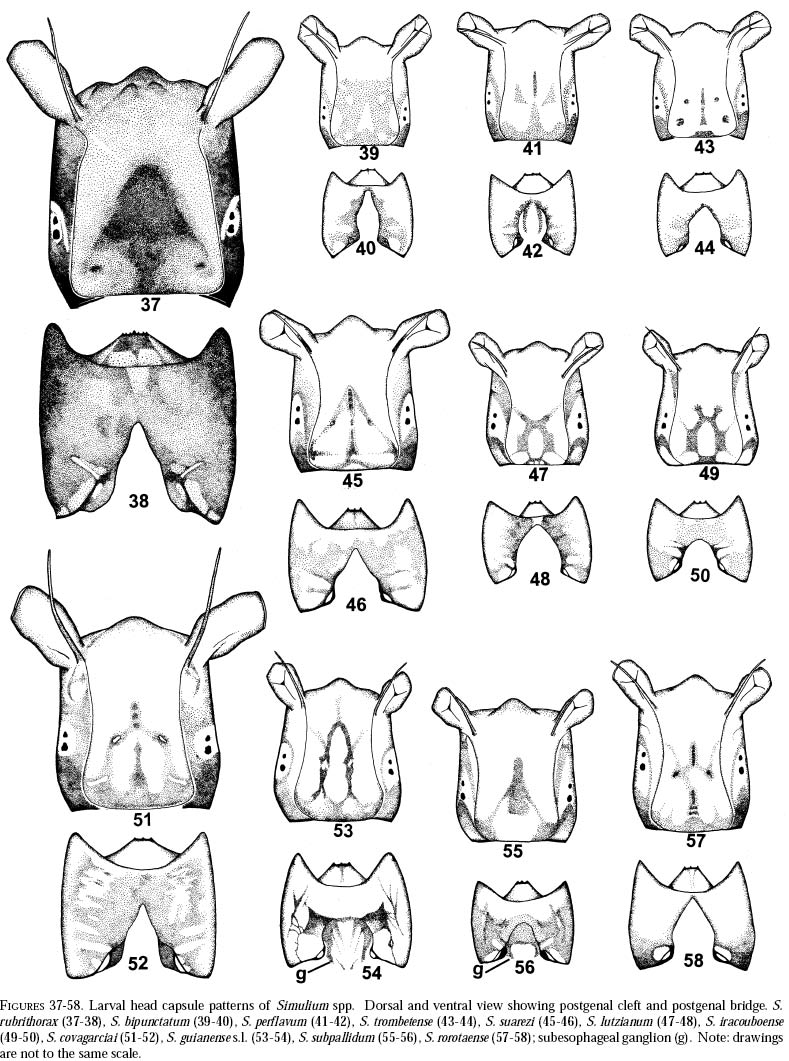
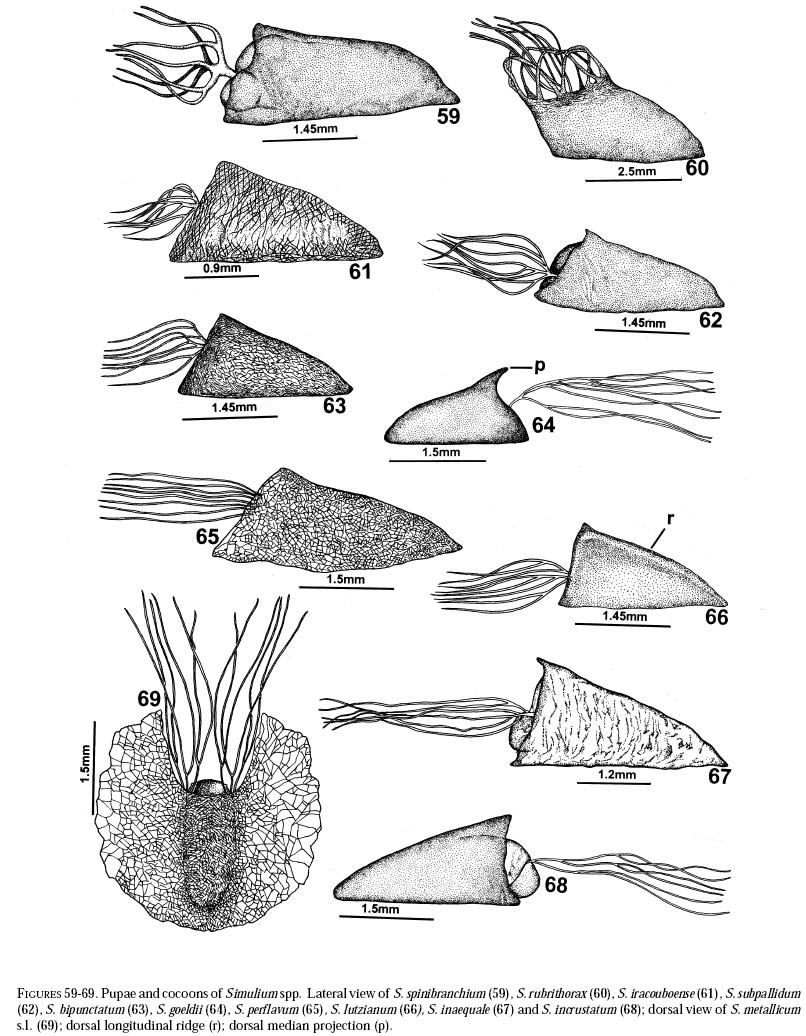
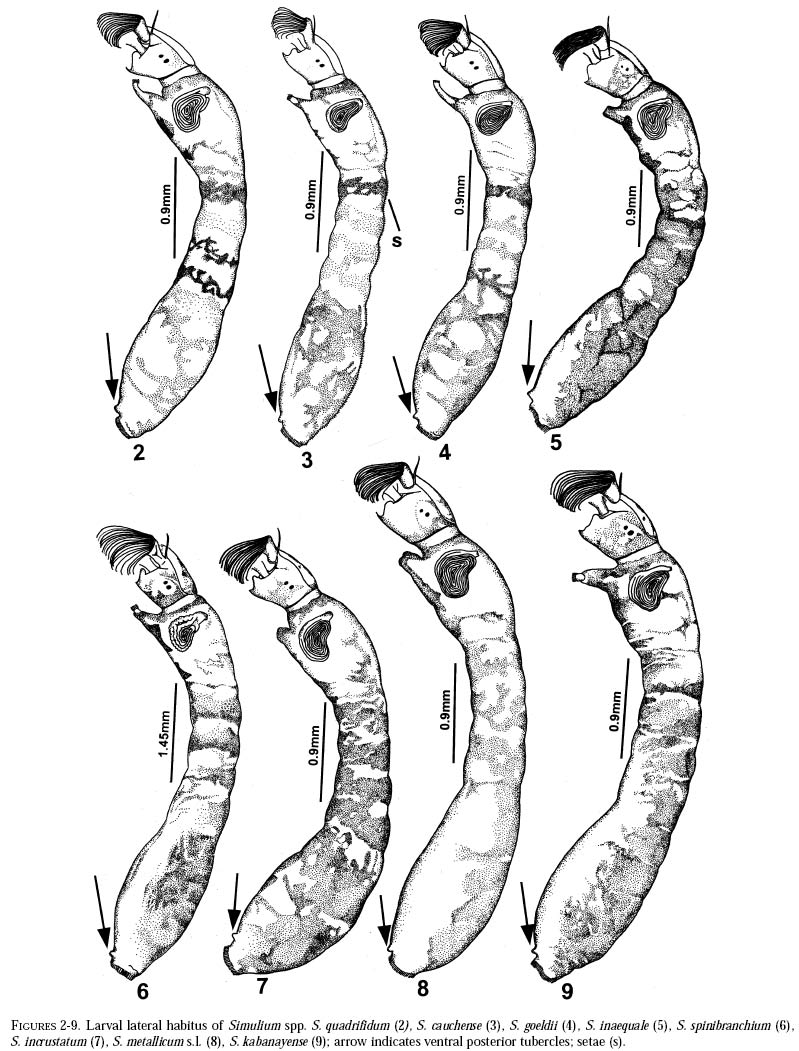
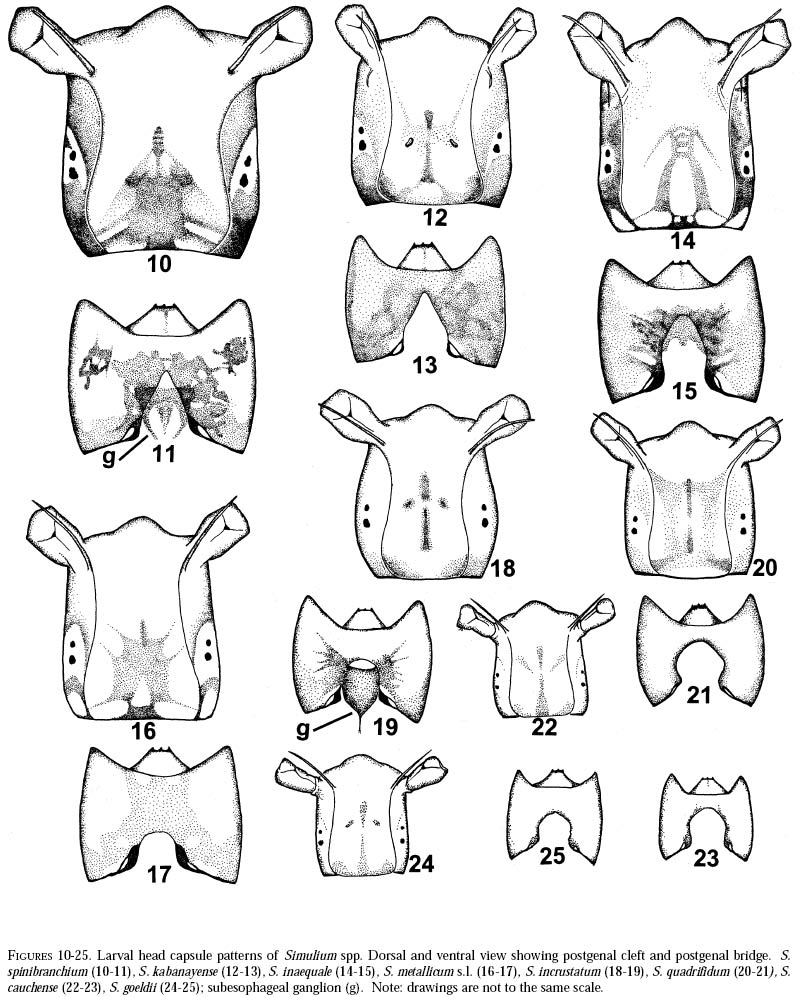
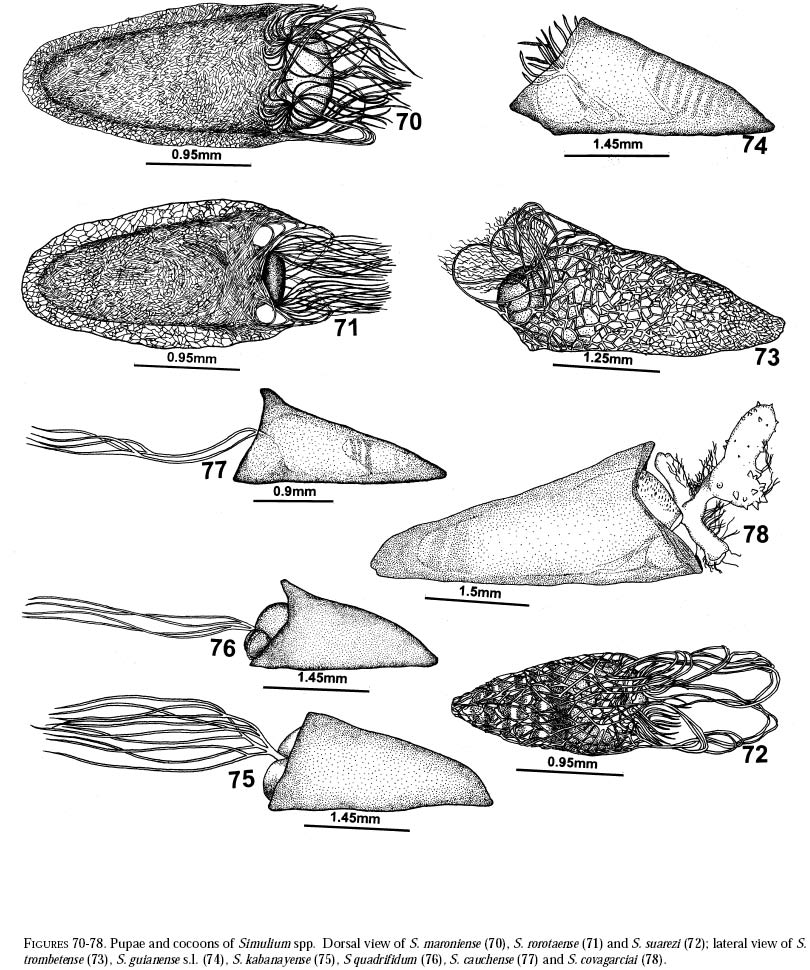
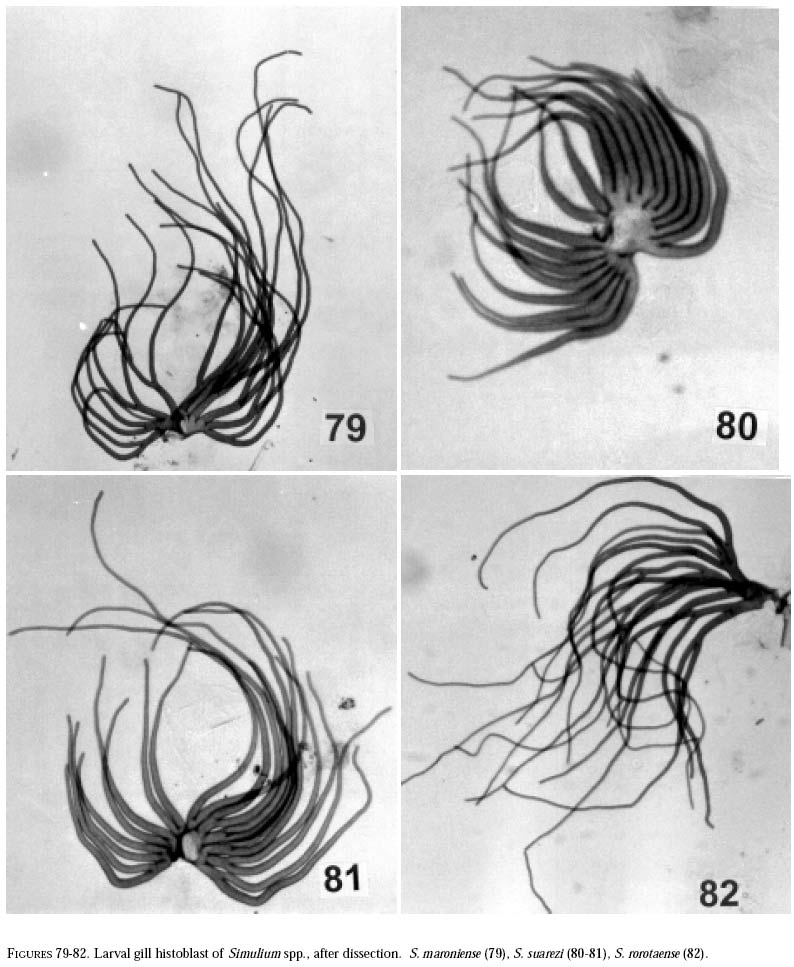
richness is still and is currently one of the taxonomically best-known underestimated. As adults, certain species of Simulium Latreille are the vectors of Onchocerca volvulus Leuckart (Nematoda: Onchocercidae), the parasite causing human onchocerciasis, a disease that has great social impact in tropical regions of Africa and the Americas. Thus, in addition to describing patter ns in streams, knowledge of simuliid diversity can provide insight into the dynamics of onchocerciasis transmission when species vectors are considered (Vivas-Martínez et al. 1998; Grillet et al. 2001). In Venezuela, taxonomic studies of black flies are particularly scarce (Briceño-Iragorry and Ortíz 1957; Ramírez-Pérez et al. 1982; Ramírez-Pérez 1983; Grillet et al. 1995; Hamada and Adler 1999), with the fauna consisting of 57 described morphospecies (Ramírez-Pérez 1983). Only one species complex (S. metallicum Bellardi cytotype E: Grillet et al. 1995) and one species group (S. perflavum Roubaud: Hamada and Adler (1999) have been studied at the cytotaxonomic level. In Brazil, a major number of people have been working on black flies taxonomy (e.g., Py-Daniel 1981, 1983; Coscarón 1990; Py-Daniel and Sampaio 1995; Shelley at al. 1997; Hamada and Adler 1998a, b, 1999). However, only 81 morphospecies (Crosskey 1988), three species complexes (Procunier 1989; Campos Gaona et al. 1996; Charalambous et al. 1996), and one species group (Hamada and Adler 1999) have been described. The species richness described so far represents about 18% (Venezuela) and 25% (Brazil) of the simuliids in the Neotropics, hence an increase in recorded species richness would be expected with an increase in taxonomic work. Little work has been done on black flies from the Gran Sabana region (Bolívar State), southern Venezuela. This region includes Canaima National Park, which is known for its high endemic species richness and numerous running-water ecosystems (Huber 1995). Seventeen species of Simuliidae have been previously reported for the area (Ramírez-Pérez 1983). Our study was intended to confirm or increase these figures, and to augment knowledge of the simuliid fauna in Venezuela and in the Neotropical Region, as well as to stimulate future systematic and ecological studies of aquatic insect communities in the region. Because the Pacaraima mountain region (Roraima,Brazil) has a long border with the Gran Sabana area and similar paleoecology ("Guayana shield", Huber 1995), we decided to include the simuliid species collected there, since both regions can share species. The work was undertaken as part of a larger research effort aimed at characterizing the taxonomy and ecology of the Amazonian simuliid fauna (Hamada and Adler 1998a,b, 1999; Hamada and McCreadie 1999). In the present paper, we present identification keys of larvae and pupae to distinguish members of the main black fly species present in the region. MATERIALS AND METHODSStudy area. The study was carried out in Canaima National Park, Gran Sabana region (Figure 1, Table 1), a high and undulating plain covering close to 30 000 km2, in the southeastern corner of Bolívar state, southern Venezuela. In addition, collections were made at the southern boundary of the Park, along the northern border of Brazil, and on the western side of the Sierra Pacaraima, Brazil (Figure 1, Table 1). Most of the Gran Sabana uplands lying between 500 and 1500 m elevation above sea level have a humid sub-montane climate, with average annual temperatures ranging between 18 and 24ºC, average annual rainfall between 2 000 and 3 000 mm, and a short dry season occurring from December to March (Huber 1995). In the extreme south of the Gran Sabana (e.g., Santa Elena de Uairén) and the Sierra Pacaraima region, the climate is of the lower montane type (Huber 1995), with average rainfall between 1 000 and 2 000 mm and a more pronounced dry season 2-4 months long from December to March or April. The area is covered mainly by treeless savannas interspersed with montane and gallery forests. It is located in the area of influence of the lower Orinoco River Basin and upper Caroní River Basin, with numerous watercourses present in the area, most of them black-water rivers (Huber 1995). Black fly sampling. The streams were chosen based on accessibility by road. Sampling was done on the main roads and trails up to km 88 along the Santa Elena de Uairén to Ciudad Guayana road in the Canaima National Park, Gran Sabana, Venezuela (Figure 1, Table 1). In Brazil, water courses were sampled along the BR-174 highway from Pacaraima village to Bananal road, in the Sierra Pacaraima (Roraima, Brazil; Figure 1, Table 1). Systematic sampling in October 1996 (rainy season) was followed by additional sampling in February (dry season) and October 1998 to supplement taxonomic studies. Larvae and pupae were collected by hand, fixed in Carnoy's solution (3 part absolute ethanol: 1 parts glacial acetic acid), and refrigerated. Pupae with pharate adults were maintained alive in plastic vials with wet filter paper to obtain the adults; after emergence they were fixed in 80% ethanol. Vouchers of all examined species are deposited in the Invertebrate Collection of Laboratorio de Biología de Vectores (MLBV), Instituto de Zoología Tropical, Universidad Central de Venezuela, Caracas, Venezuela and in the Invertebrate Collection of the Instituto Nacional de Pesquisas da Amazônia (INPA), Manaus, Amazonas, Brazil. RESULTS AND DISCUSSIONTwenty-one simuliid species were collected in the immature stages, from a total of 45 (Table 1). In Gran Sabana the following species of Simulium were collected: S. bipunctatum Malloch (Figures 31, 39, 40, 63), S. cauchense Floch & Abonnenc (Figures 3, 22, 23, 77), S. guianense s.l. Wise (Figures 36, 53, 54, 74), S. goeldii Cerqueira & Nunes de Mello (Figures. 4, 24, 25, 64), S. ignacioi Ramírez-Pérez & Vulcano, S. inaequale Paterson & Shannon (Figures 5, 14, 15, 67), S. incrustatum Lutz (Figures 7, 18, 19, 68), S. iracouboense Floch & Abonnenc (Figures 27, 49, 50, 61), S. kabanayense Ramírez-Pérez & Vulcano (Figures 9, 12, 13, 75), S. lutzianum Pinto (Figures 26, 47, 48,66), S. maroniense Floch & Abonnenc (Figures 70, 79), S. metallicum s.l. (Figures 8, 16, 17, 69), S. perflavum (Figures 30, 41, 42, 65), S. quadrifidum Lutz (Figures 2, 20, 21, 76), S. spinibranchium Lutz (Figures 6, 10, 11, 59), S. suarezi Ramírez-Pérez, Rassi, Ramírez (Figures 28, 45, 46, 72, 80, 81) and S. subpallidum Lutz (Figures 32, 55, 56, 62). In the Pacaraima mountains the following species of Simulium were collected: S. cauchense, S. covagarciai Ramírez-Pérez, Yarzábal, Takaoka, Tada & Ramírez (Figures 33, 51, 52, 78), S. lutzianum, S. maroniense, S. metallicum s.l., S. perflavum, S. rorotaense Floch & Abonnenc (Figures 29, 57, 58, 71, 82), S. rubrithorax Lutz (Figures 35, 37, 38, 60) and S. trombetense Hamada, Py-Daniel & Adler (Figures 34, 43, 44, 73). Simulium ignacioi was synonymized with S. rorotaense by Shelley et al. (1984, 1997); however, Hamada and Adler (1998a) considered them as distinct species based on gill filament numbers and chromosomal configuration (Hamada N, Adler PH, Grillet ME, unpublished data). This is the first report of S. covagarciai for the study area and for Brazil. This species was described from Sierra Parima by Ramírez-Pérez et al. (1984). Similarly, this is the first report of a species in the S. metallicum complex in southern Venezuela (southern region of the Orinoco River Basin) and Brazil. Ramírez-Pérez et al. (1977) described S. morae from Sierra Parima and, by the description of this species, we are certain that it belongs to the S. metallicum complex. This nominal species comprises at least 12 cytotypes (Conn et al. 1989; Arteagas and Muños de Hoyos 1999). Cytological studies will be necessary to define the cytotypes present in the study area, as well as their biting behavior since S. metallicum cytotype E is associated with onchocerciasis transmission in northern Venezuela (Grillet et al. 1995). Likewise, S. guianense s.l. and S. incrustatum were collected in the Gran Sabana. These species play an important role in onchocerciasis transmission in the Amazonian focus of southern Venezuela and northern Brazil (Basáñez et al. 1988; Shelley et al. 1997; Grillet et al. 2000), and consequently should be the aim of future cytotaxonomic and entomological studies in the region. This work presents the first simuliid taxonomic keys based on larval and pupal characters in Venezuela, and provides, along with the keys of Ramírez-Pérez (1983) to adults and pupae, a more comprehensive entomological baseline for new systematic and ecological studies planned in the Gran Sabana area. With further samples, especially from the higher plateau or "tepuyes" of the Canaima National Park, and other habitats not incorporated in this study, the described simuliid richness of this region will certainly increase. Identification key to last-instar larvae 1 Paired ventrolateral posterior tubercles present (Figures 2-9) ..................................................... 2 -Paired ventrolateral posterior tubercles absent (Figures 26-36) .................................................. 9 2[1] Subesophageal ganglion pigmented (Figures 11,19) ........................................................................ 3 - Subesophageal ganglion not pigmented .............. 4 3[2] Gill filaments in situ (Figure 6) thick, dissected with 8 filaments .................................. S. spinibranchium -Gill filaments in situ (Figure 7) thin, dissected with 6 filaments ....................................... S. incrustatum 4[2] Distal portion of postgenal cleft rounded (Figures 17, 21, 23, 25) ........................................................ 5 -Distal portion of postgenal cleft pointed (Figures 13, 15) ................................................................... 8 5[4] Postgenal bridge large, more than 11/2 imes longer than postgenal cleft length (Figure 17); head capsule pigmented dorsally with clear spot on basal half (Figure 16) ................................. S. metallicum s.l. -Postgenal bridge smaller than the length of postgenal cleft (Figures 21, 23, 25); head capsule dorsally without clear spot on basal half (Figures 20, 22, 24) ........................................................................ 6 6[5] Postgenal cleft not wide at the apical portion (Figure 25); lateral mandibular process bifid or trifid; second antennal article larger than the first; dissected gill histoblast with 8 filaments ...................... S. goeldii - Postgenal cleft wide at the apical portion (Figures 21, 23); lateral mandibular process single; second antennalarticle smaller than the first; dissected gill histoblast with 4 filaments ................................. 7 7[6] Body surrounded by visible setae (Figure 3); dissected gill histoblast with filaments branching far away from the base (Figure 77) ........................... S. cauchense -Body not surrounded by visible setae (Figure 2); dissected gill histoblast with filaments branching near the base (Figure 76) ....................... S. quadrifidum 8[4] Head capsule dorsally with dark pattern, as in figure
14; dissected gill histoblast with 6 filaments .......... ........................................................S.
inaequale 9[2] Subesophageal ganglion pigmented (Figures 54, 56) ...................................................................... 10 - Subesophageal ganglion not pigmented ............ 11 10[9] Body surrounded by visible setae (Figure 36); head capsule as in Figure 53; dissected gill histoblast with 12 filaments ................................. S. guianense s.l. -Body not surrounded by visible setae; head capsule as in Fig. 55; dissected gill histoblast with 8 filaments ..................................................... S. subpallidum 11[9] Head capsule heavily pigmented (Figure
37); large larvae (> 10 mm); body wider at posterior region getting
narrower anteriorly, as in figure
35 ............ ...................................................... S.
rubrithorax 12[11] Gill histoblast in situ large, as in Fig. 33; dissected gill histoblast with bulbous expansions (Figure 78) ..... ........................................................... S. covagarciai -Gill histoblast in situ not large; dissected gill histoblast without bulbous expansions ............................. 13 13[12] Head capsule dorsally with a dark ring-shaped pattern, with oval clear spot (Figures 47, 49) .... 14 -Head capsule dorsally without a dark ring-shaped pattern and oval clear spot ............................... 15 14[13] Postgenal bridge small, less than 20% of postgenal cleft length (Figure 48) ...................... S. lutzianum -Postgenal bridge larger, at least same size as postgenal cleft length (Figure 50) .................. S. iracouboense 15[13] Head capsule with 5 clear spots on a dark background at dorsal median
region (Figure 39); postgenal
cleft as in Figure 40 .............................. S.
bipunctatum 16[15] Body usually with a white dorso-lateral stripe on last third of body (Figure 28); dissected gill histoblast usually with 20 clear filaments with enlarged basal portion, as in Figures 80, 81 .................. S. suarezi -Body usually without a white dorso-lateral stripe on last third of body (Figures 29, 30, 34); dissected gill histoblast with 17-23 darker filaments, with basal portion not enlarged, as in Figures 79, 82 ......... 17 17[16] Gill histoblast in situ with numerous thin filaments coming out from thick trunk, dissected with more than 100 filaments (Figure 73) ........ S. trombetense -Gill histoblast in situ not with numerous thin filaments, basal trunk not as thick as above, dissected with less than 25 filaments ............................................ 18 18[17] Gill histoblast dissected with 8 filaments (Figure 65) postgenal cleft as in Figure 42 ........... S. perflavum -Gill histoblast dissected with 14-23 filaments (Figures 79, 82); postgenal cleft as in Figure 58 .............. 19 19[18] Gill histoblast dissected with 14-17 filaments (usually 16) ..................................................... S. ignacioi -Gill histoblast dissected with 17-23 filaments (usually 18-21) ............................................................. 20 20[19] Dissected gill histoblast with thick, dark filaments branching near the
base, with dorsal filaments shorter than ventral ones; in anterior view, filaments
forming open rosette basally (Figure
79) ....... S. maroniense Identification Key to Pupae 1 Gill without bulbous expansions ....................... 2 2[1] Gill with 4 filaments ......................................... 3 3[2] Gill filaments branching near base (Figure
76) ...... .................................................... S.
quadrifidum 4[2] Gill with 6 filaments ......................................... 5 5[4] Gill filaments branching near base (Figures
67, 69) ........................................................................
6 6[5] Cocoon with closed mesh, without lateral expansions (Figure
67) ........................................ S.
inaequale 7[4] Gill with 8 filaments (Figures
61, 62, 65) ............ 8 8[7] Cocoon boot shaped (Figure
60) ...... S. rubrithorax 9[8] Cocoon with dorsal median projection; gill filaments branching at different
distances from base (Figure
64) ............................................................ S.
goeldii 10[9] Gill thick basally, with rigid branches (Figure
59) . ................................................ S.
spinibranchium 11[10] Cocoon with dorsal median longitudinal ridge (Figure
66) ................................................... S.
lutzianum 12[11] Gill filaments branching from one common trunk, usually, at same heights (Figure 65) .... S. perflavum -Gill filaments branching at different heights near base (Figures 61, 62), main trunk branching in three primary branches ............................................ 13 13[12] Cephalic trichomes short and simple ............................................................................. S. iracouboense -Cephalic trichomes bifid or trifid ..................... 14 14[13] Head capsule and thorax dark in color, frontoclypeus with no or few platelets
.................. S. subpallidum 15[7] Gill filaments with spinose apex (Figure
74) ............................................................ S.
guianense s.l. 16[15] Gill with 10-17 filaments ................................. 17 17[16]Gill with 10-14 filaments; cocoon without lateral opening (Figure
75)
....................... S. kabanayense 18[16] Gill with 17-23 filaments ................................. 19
19[18] Gill with 20 filaments, branching near an enlarged central base; filaments enlarged basally as in Figures 72, 80, 81 .............................................. S. suarezi -Gill with 17-23 filaments (Figures 70, 71), not branching near an enlarged central base, filaments not enlarged basally, as in Figures 79, 82 .......... 20 20[19] Gill with thin, lightly pigmented filaments, varying in size and branching pattern; in anterior view, filaments projected forward, not forming rosette (Figure 82) ........................................ S. rorotaense -Gill with thick, darkly pigmented filaments; dorsal filaments shorter than ventral ones; in anterior view, filaments forming open rosette basally (Figure 79) ...................................................... S. maroniense ACKNOWLEDGEMENTSWe thank John W. McCreadie, Christian Steiner, César Delgado, Yamile B. Alencar, Sílvio J. Reis and Claudia Ríos-Velásquez who assisted us in the field. INPARQUES (Venezuela) granted us the opportunity to collect black flies in Canaima National Park. PPI-1/3400 (INPA) and CNPq (201165/93-7) provided partial financial support to N. Hamada, whereas M. E. Grillet was financed by CDCH-UCV (1996, 1997). Deyse Queiróz made the larval and pupal habitus drawings, and Jorge Dácio inked the map. Philip M. Fearnside reviewed the manuscript. REFERENCES
The following images related to this document are available:Photo images[em01003f10-25.jpg] [em01003f79-82.jpg] [em01003f37-58.jpg] [em01003f26-36.jpg] [em01003f59-69.jpg] [em01003f2-9.jpg] [em01003f70-78.jpg] [em01003f1.jpg] [em01003t1.jpg] |
| |||||||||
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}