 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
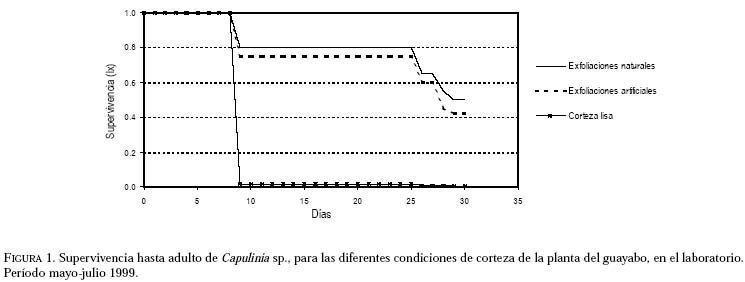
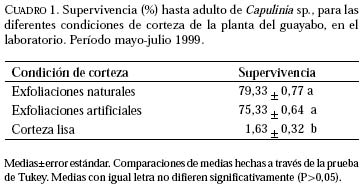
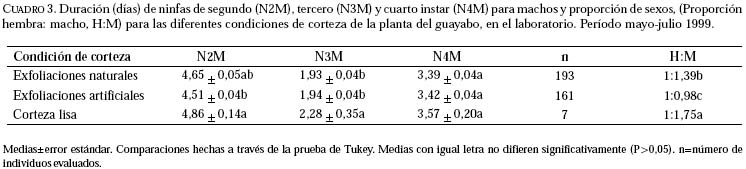
Entomotropica, Vol. 16, No. 1, Abril/April, 2001, pp. 21-27 Efecto físico de las exfoliaciones de la corteza del guayabo (Psidium guajava) sobre Capulinia sp. cercana a jaboticabae von Ihering (Hemiptera: Eriococcidae)Francis Geraud-Pouey, Dorys T. Chirinos, Gustavo Romay Laboratorio de Manejo Integrado de Plagas en Frutales y Hortalizas, Unidad Técnica Fitosanitaria, Facultad de Agronomía, La Universidad del Zulia. Telefax: (58 61) 597113. E-mail: fgeraud@luz.ve, fgeraudp@hotmail.com, dtchirinos@hotmail.com, gustavoromay@hotmail.com Code Number: em01007 RESUMEN GERAUD-POUEY F, CHIRINOS DT, ROMAY G. 2001. Efecto físico de las exfoliaciones de la corteza del guayabo sobre Capulinia sp. cercana a jaboticabae von Ihering (Hemiptera: Eriococcidae). Entomotropica Vol. 16(1):21-27. Desde su aparición en Venezuela a principios de 1993, la mota blanca del guayabo, Capulinia sp. ha constituido el problema entomológico de mayor relevancia en guayabo, Psidium guajava L. Sin embargo, en el campo se observa que plantas con corteza lisa (con pocas o sin exfoliaciones) son menos infestadas por este insecto. En consecuencia, durante el período mayo-julio de 1999 se evaluó en el laboratorio el efecto físico de las exfoliaciones de la corteza del guayabo sobre el establecimiento, la supervivencia y el desarrollo de Capulinia sp. Fueron incluidas tres condiciones de planta hospedera (selección local Criolla Roja): con exfoliaciones naturales, con corteza mecánicamente alisada (corteza lisa) y exfoliaciones sustituidas por pedazos de papel absorbente adherido a la corteza alisada (exfoliaciones artificiales). La supervivencia de las ninfas de primer instar fue significativamente incrementada (>70%, P≤ 0,05) por las exfoliaciones (naturales y artificiales) como sitios debajo de los cuales se establecen para alimentarse. No obstante, la duración del desarrollo fue poco afectada por la condición física de la corteza. La corteza lisa podría constituir una fuente de resistencia del guayabo para disminuir la incidencia y daños por Capulinia sp. en ese frutal. Palabras clave adicionales: Coccoidea, escamas, guayaba, mota blanca del guayabo, planta hospedera. ABSTRACT GERAUD-POUEY F, CHIRINOS DT, ROMAY G. 2001. Physical effect of exfoliation of guava tree bark on Capulinia sp. near to jaboticabae von Ihering (Hemiptera: Eriococcidae). Entomotropica Vol 16(1):21-27. Since it appeared in Venezuela in early 1993 the guava cottony scale, Capulinia sp. has become the greatest entomological problem on guava trees, Psidium guajava L. However in the field, trees with smooth bark (with little or without exfoliations) tend to be much less infested by this insect. Therefore, during May-July 1999 the physical effect of bark exfoliations on the establishment, survival and development of Capulinia sp. was assessed, in the laboratory. Three host plant /Criolla oja local selecction) conditions were included: with natural exfoliations, with bark mechanically smoothed (smooth bark) and exfoliations substituted by pieces of absorbent paper adhered to the smoothed bark (artificial exfoliations). Survival of first instar nymphs (crawlers) was significantly increased (>70%, P≤ 0.05) by the presence of exfoliations (natural and artificial) as places under which they settle down to feed. However, duration of development was little affected by the existence of exfoliations. Bark smoothness could constitute a source of resistance of guava plants to diminish the incidence and damages by Capulinia sp. Additional key words: Coccoidea, guava, guava cottony scale, host plant. INTRODUCCIÓN En Venezuela el cultivo del guayabo, Psidium guajava L. se incrementó vertiginosamente a partir de comienzos de la década de 1980 (Geraud-Pouey 2000) llegándose a estimar la existencia de unas 5000 ha de huertos, el 80% de éstos en el estado Zulia (Araujo et al. 1997). Desde sus comienzos el cultivo de ese frutal ha sido afectado por problemas fitosanitarios, en buena parte favorecidos por el acelerado incremento en la superficie sembrada aunado al deficiente soporte científico-tecnológico para enfretarlos. Así, durante 1990-1995 se perdieron aproximadamente 2000 ha de huertos a consecuencia de muerte regresiva de árboles asociada a nemátodos agalladores de raíces Meloidogyne sp. y hongos de suelo, complementada por los daños causados por la mota blanca del guayabo, Capulinia sp. Esta última fue detectada en el país en 1993 (Cermeli y Geraud-Pouey 1997) y constituye un nuevo registro para la ciencia (Chirinos 2000). El insecto forma sus colonias principalmente sobre tallos y ramas del árbol. Las fuertes infestaciones producidas por el alto potencial reproductivo de este insecto (Chirinos et al. 1997a, b; Chirinos 2000) permitido por la entonces inexistencia de enemigos naturales específicos y efectivos, facilitó su dispersión en casi todo el territorio nacional. En 1996 fue detectado un parasitoide más específico de este insecto fitófago (Geraud-Pouey et al. 1997; Cermeli y Geraud-Pouey 1997). Dicho parasitoide fue recientemente identificado por el Dr. John Noyes (British Natural Histor y Museum) como Metaphycus sp. (muy probablemente especie no descrita) (Chirinos-Torres et al. 2000). Aunque a partir de la aparición del parasitoide, las infestaciones por Capulinia sp. disminuyeron considerablemente, ésta no deja de ser problema para la producción de guayaba (Cermeli y Geraud-Pouey 1997; Geraud-Pouey y Chirinos 1999; Chirinos-Torres et al. 2000). Para comprender mejor los problemas entomológicos causados por Capulinia sp. en huertos de guayabo es necesario conocer detalles de la relación insectoplanta. Generalmente las ninfas de primer instar de Capulinia sp. se establecen debajo de exfoliaciones de la corteza del guayabo (Cermeli y Geraud-Pouey 1997). En el campo hemos observado que plantas con menos exfoliaciones de corteza suelen tener menores infestaciones por este insecto. Además, durante la fase preliminar de estudios de biología en 1996, para facilitar las observaciones de Capulinia sp., a las primeras cuatro plantas pequeñas utilizadas (sembradas en macetas); les fueron eliminadas mecánicamente las exfoliaciones de corteza, previo a la colocación de los huevos del insecto (inicio de cohorte). En ninguna de esas plantas se establecieron las ninfas de primer instar. Algunas fueron encontradas muertas, pero la mayoría desapareció. Probablemente deambularon hasta perderse en el suelo, en la base de la planta. Todo esto sugiere que las exfoliaciones de la corteza del guayabo juegan un papel importante en el establecimiento del insecto, para comenzar a alimentarse de la planta y desarrollar colonias. Con el fin de aclarar esta situación, se llevó a cabo un experimento de laboratorio para evaluar el efecto físico de las exfoliaciones de corteza del guayabo sobre el establecimiento, la supervivencia y desarrollo de la mota blanca del guayabo, Capulinia sp. MATERIALES Y MÉTODOS La investigación se realizó durante el período mayo-julio 1999 en el Laboratorio de Manejo Integrado de Plagas en Fr utales y Hortalizas (temperatura: 26,74+1,50°C, humedad relativa 82,47+6,77%), Unidad Técnica Fitosanitaria, Facultad de Agronomía, La Universidad del Zulia, Maracaibo, Venezuela. Las plantas experimentales fueron mantenidas bajo paneles de iluminación (125 x 45 cm, largo x ancho) con cuatro tubos fluorescentes de 40 w c/u y cinco bulbos incandescentes de 60 w c/u, con una duración de 12 horas luz, sin excluir la luz natural. Plantas experimentales. Fueron utilizadas plantas de guayabo "Criolla Roja", una selección local comúnmente sembrada en la planicie de Maracaibo, cuya corteza produce abundantes exfoliaciones, lo cual la hace muy adecuada para el desarrollo de colonias de Capulinia sp. Parte de estas plantas sirvió para mantener la colonia del insecto bajo estudio y otras fueron utilizadas como sustrato experimental. Las plantas para mantenimiento de colonias fueron cultivadas en bolsas de polietileno de aproximadamente 2 kg de capacidad de mezcla de suelo (dos partes de arena limosa y una parte de materia orgánica vegetal), dejadas al aire libre y fertilizadas semanalmente con 5 g/planta de una fórmula completa (12-12-17). Las plantas destinadas para sustrato experimental inicialmente fueron sembradas y tratadas siguiendo el procedimiento indicado para el mantenimiento de colonias. Cuando tenían alrededor de cuatro meses de edad y la base de su tallo había alcanzado aproximadamente 1 cm de diámetro, cada una era transplantada a un envase de plástico de 10,5 x 9 cm (altura x diámetro) y capacidad aproximada de 0,5 kg de suelo. Cada envase tenía tapa enroscada, la cual se colocaba con el fin de evitar la pérdida del suelo durante las observaciones al estereoscopio. Dicha tapa tenía un agujero central de 1,5 cm de diámetro, para alojar la base del tallo de la planta, la cual era introducida mediante una abertura radial hasta el borde externo de la misma. La tapa tenía cuatro orificios equidistantes, de 0,5 cm de diámetro, utilizados para regar la planta. En la parte inferior de la pared del envase, también había cuatro orificios equidistantes para drenar el exceso de agua de riego. Previo al trasplante, la raíz de fijación era podada para facilitar el acomodo de las raíces de absorción dentro del envase. La parte aérea de la planta también era podada, dejando un tallo recto de aproximadamente 25 a 30 cm de altura con dos a cuatro hojas funcionales en el tope. Posterior al transplante, las plantas fueron mantenidas por dos semanas en recuperación antes de ser utilizadas en los experimentos. Colonias de laboratorio. Con el fin de disponer de huevos de Capulinia sp. para los experimentos, fueron mantenidas colonias con plantas de vivero naturalmente infestadas y luego introducidas al laboratorio. Las plantas eran mantenidas dentro de jaulas entomológicas de madera ( 32 x 37 x 29 cm largo x ancho x alto) con una manga de tela en la puerta frontal, tope de vidrio en bisel y parte posterior cerrada con tela de organza, modificada después de Peterson (1945). Experimentos. Las plantas experimentales infestadas con huevos de Capulina sp. también eran mantenidas dentro del mismo tipo de jaulas entomológicas utilizadas para mantenimiento de colonias. Las observaciones fueron realizadas bajo un microscopio estereoscópico (amplitud de aumento: 6,3-40×). Al momento de la observación, la planta era colocada horizontal, apoyando el envase sobre una base de poliuretano (15×15×3 cm, ancho×largo×grosor), con una acanaladura de sección semicircular, sobre el cual la planta era rotada para observar los insectos distribuidos alrededor de su tallo. Los desplazamientos longitudinales se lograban moviendo la base en ese sentido. Para el inicio de los experimentos a hembras de la colonia se les retiraron las masas de huevos existentes con un pincel fino. Esas hembras eran marcadas con un alfiler entomológico clavado cerca de ellas, sobre el tallo de la planta y se les dejaba oviponer por 20 horas, lo cual aseguraba aproximadamente 50-130 huevos/hembra. Al cabo de ese tiempo, los huevos eran retirados con el pincel y colocados sobre la planta utilizada como sustrato experimental, aproximadamente a media altura del tallo. Se incluyeron plantas con tres condiciones de corteza a manera de tratamientos, la unidad experimental consistió de una planta y se realizaron seis repeticiones. En uno de los tratamientos, a las plantas se les dejó intacta la corteza (con exfoliaciones naturales), mientras que en los otros dos les fueron eliminadas mecánicamente, con una pinza. En uno de estos últimos, las exfoliaciones fueron sustituidas por pedazos rectangulares (1 x 0,5 cm, largo x ancho) de papel absorbente, tipo toallín, humedecido para asegurar su adherencia a la corteza. Al hacerlo, eran colocados longitudinalmente, adhiriendo el borde corto superior y dejando parcialmente levantado el resto, de modo de simular las exfoliaciones naturales al secarse el papel. A las plantas del tercer tratamiento, simplemente se les dejó la corteza lisa. Posteriormente a su colocación sobre las plantas experimentales, los huevos fueron observados diariamente al microscopio estereoscópico hasta la eclosión. Después de la eclosión se esperaba durante dos días para que las ninfas se fijaran al tallo introduciendo el estilete bucal. Una vez fijadas, para facilitar las observaciones, las exfoliaciones (naturales y artificiales) eran retiradas con una pinza. En el caso de las ninfas macho, cuando estaban próximas a completar el segundo instar (aproximadamente dos días antes), sobre plantas con exfoliaciones (naturales y artificiales), se colocaban en el tallo pedazos de papel absorbente de la forma antes descrita, debido a que dichas ninfas buscan exfoliaciones y hembras cercanas para formar sus capullos dentro de los cuales se suceden los instares posteriores (Cermeli y Geraud-Pouey 1997; Chirinos et al. 1997a, b; Geraud-Pouey y Chirinos 1999; Chirinos 2000). Algunos machos formaron sus capullos sobre el tallo, pero otros lo hicieron sobre el papel absorbente o sobre los brotes foliares. En estos casos, una vez formado el capullo, los pedazos de brotes y/o de toallín eran retirados y se colocaban individualmente dentro de cápsulas de gelatina transparente (Lilly No. 1) donde se continuaban las observaciones hasta la salida del insecto adulto. La evaluación del efecto físico de las exfoliaciones de corteza consideró supervivencia hasta adulto, duración de huevo, de fases juveniles y proporción de sexos. Los diferentes instares ninfales por los cuales pasan hembras y machos en esta especie, han sido previamente referidos y descritos (Geraud-Pouey y Chirinos 1999; Chirinos 2000). La supervivencia y la proporción de sexos fueron analizadas con un diseño de bloques al azar para lo cual previamente fueron transformadas a la función arcoseno (x)1/2 para ajustarlas a la distribución normal. La duración de instares fue analizada mediante un diseño en parcelas divididas, donde la parcela principal correspondió a los individuos evaluados y la subparcela a los tratamientos. Los análisis estadísticos fueron hechos mediante el Modelo Lineal General (GLM). Todas las medias fueron comparadas a través de la prueba de Tukey (P≤0,05). Los análisis se realizaron con el programa estadístico SAS (SAS Institute 1996). RESULTADOS Y DISCUSIONSupervivencia. La Figura 1 muestran las curvas de supervivencia de Capulinia sp. para plantas con las diferentes condiciones de corteza y los resultados de comparaciones estadísticas se presentan en el Cuadro 1. Allí se observa que la misma fue significativamente inferior sobre plantas con cortezas lisas, con un valor promedio de 1,63%, y una amplitud de variación entre 0 y 6,43% entre repeticiones. Además es importante señalar que la supervivencia bajo esta condición fue consecuencia del establecimiento de ninfas de primer instar en algunas leves exfoliaciones de corteza remanentes de la eliminación mecánica que se hizo a estas plantas o de algún accidente físico de la misma, tales como cicatrices de brotes u hojas eliminadas para alisar el tallo. Algunos investigadores han reportado diferencias en la supervivencia de otras especies de artrópodos sobre diferentes especies de plantas, así como cultivares o genotipos de la misma especie de plantas hospederas, debido a factores asociados con la nutrición (Narváez y Notz 1993; Jordano y Gomariz 1994; Boavida y Neuenschwander 1995; Soria y Mollena 1995) o ligado a sustancias químicas de la planta hospedera (Yu y Terriere 1974; Kennedy y Yamamoto 1979; Isman y Duffey 1982; Reyes 1986; Romanow et al. 1991; Broadway y Villani 1995; Soria y Mollema 1995; Dominguez-Gil y McPheron 1997; Geraud-Pouey y Chirinos 1999; Chirinos 2000). No obstante, en este caso las diferencias de supervivencia se deben al efecto físico de la corteza de las plantas. Los factores físicos de la planta pueden afectar el desarrollo y supervivencia de los artrópodos fitófagos. Ha sido referida mortalidad en varias especies de artrópodos al ser atrapados por los exudados producidos por tricomas glandulares (Johnson 1956; Gentile et al. 1968, 1969; Gibson 1974; Gibson y Turner 1977). La densa pubescencia de plantas de soya interfiere con la oviposición de algunas especies de saltahojas (Painter 1951; Kogan 1975) mientras que en plantas de algodón la pubescencia favorece la oviposición de Heliothis spp. (Stadelbacher y Scales 1973). Los factores físicos también pueden afectar la supervivencia de enemigos naturales interfiriendo así con su efectividad para regular poblaciones de artrópodos fitófagos (Geraud 1986; Robertson y Hoffman 1989). Los resultados obtenidos en cuanto a supervivencia corroboran experimentalmente que en esta especie de Capulinia, las ninfas de primer instar muestran una marcada preferencia por las exfoliaciones de corteza, como sitio bajo los cuales fijarse para su posterior desarrollo. De hecho, cuando fueron colocados los pedazos de papel absorbente en el tallo para simular las exfoliaciones naturales previamente eliminadas en forma mecánica, Capulinia sp. también se estableció y aunque la supervivencia fue inferior que en las plantas con corteza natural (Figura 1), esas diferencias no resultaron estadísticamente significativas (Cuadro 1). En este ensayo, el total de la mortalidad ocurrió en ninfas de primer instar para todas las condiciones evaluadas, debido a fallas en mayor o en menor grado en el establecimiento del insecto sobre la planta. Esto corrobora lo observado en trabajos previos (Chirinos et al. 1997a, b; Chirinos 2000). Este no parece ser el único caso donde el establecimiento de los individuos durante el inicio de su desarrollo resulta crítico para sobrevivir. Prezsler y Price (1988) observaron que el factor más importante de mortalidad en Euura lasiolepis (Hymenoptera: Tenthredinidae) fue la muerte de larvas Duración del desarrollo. El Cuadro 2 muestra la duración (en días) de la fase de huevo así como de ninfas de primer instar y segundo instar hembras. Para la fase de huevo no se detectaron diferencias, probablemente porque los huevos provenían de la colonia y además las masas de huevos fueron colocadas sobre las plantas sin ser influenciadas por las exfoliaciones de la corteza. Aunque no resultó significativa, la duración de las ninfas de primer instar, fue ligeramente mayor en plantas con cortezas lisas. La ninfa de primer instar es la fase de dispersión, en la cual a partir de su fijación el insecto comienza a alimentarse y se vuelve sedentario. Probablemente, la mayor duración de las ninfas de primer instar sobre corteza lisa tenga que ver con el mayor tiempo de búsqueda de un lugar adecuado para fijarse. En la fase sedentaria, o ninfas hembras de segundo instar, las duraciones se uniformizaron sobre todas las plantas con las tres condiciones evaluadas. En el caso de los machos (Cuadro 3), la duración del segundo instar sobre plantas bajo las tres condiciones de corteza varió entre 4,51-4,86 (7,76 %). En plantas con cortezas lisas la duración fue significativamente mayor que en plantas con exfoliaciones artificiales (P≤0,05). Las diferencias encontradas en la duración del segundo instar del macho, sugiere que la ausencia de exfoliaciones, al igual que en el primer instar, obliga a las ninfas a movilizarse más hasta encontrar un sitio adecuado para fijarse y/o formar el capullo, lo cual podría alargar el tiempo de desarrollo de esas fases. En el macho las ninfas de segundo instar mantienen su movilidad. En el caso de las ninfas de tercer instar, la duración fue mayor en plantas con cortezas lisas (18,13 %; P≤ 0,05 ). Para cuarto instar, también resultó ligeramente mayor (5,31 %), pero no significativa. En vista que el tercer y cuarto instar ocurren dentro del capullo, sin alimentarse ni desplazarse, es de suponer que esas diferencias son producto de la relación insecto planta durante los instares previos. En general, aunque las diferencias resultaron consistentes, la duración del desarrollo parece ser poco afectada por la existencia de exfoliaciones (4,85 % de diferencia, para machos). Han sido reportados casos de otros insectos donde la duración del desarrollo no se ve afectada por la planta hospedera (Boavida y Neuenschwander 1995, Sánchez et al. 1997), aunque ésta influya de alguna otra manera en la biología de los mismos. Proporción de sexos. Con respecto a la proporción de sexos (Cuadro 3), fue superior la proporción de machos en plantas con cortezas lisas. Dadas las marcadas variaciones en la proporción de sexos de esta especie (Chirinos 2000), es difícil medir el efecto de la condición de la corteza sobre esta variable. Las diferencias en número de individuos evaluados en las diferentes condiciones corteza de plantas es consecuencia de las variaciones en la supervivencia bajo las mismas, lo cual fue tratado previamente. El marcado efecto de las características físicas de la corteza sobre la supervivencia de las ninfas de primer instar de Capulinia sp., podría ser utilizado como una alternativa para disminuir las infestaciones por este insecto en huertos de guayabo. Dada la importancia del mismo como plaga del guayabo en Venezuela, la corteza lisa podría constituir un factor de resistencia al insecto, susceptible de ser utilizado, mediante manipulaciones genéticas, en programas de manejo integrado de plagas de este frutal (Geraud-Pouey y Chirinos 1999). AGRADECIMIENTOSAl Consejo Nacional de Investigaciones Científicas y Tecnológicas (CONICIT) por haber subvencionado los estudios de Maestría de la Ing. Agr. Dorys T. Chirinos, dentro de los cuales se realizó esta investigación y al Consejo de Desarrollo Científico y Humanístico (CONDES) de LUZ (Proyecto 2154-95), por haber financiado parcialmente dicha investigación. Así mismo, al Centro Frutícola-CORPOZULIA quien dono las plantas utilizadas en los experimentos. REFERENCIAS
The following images related to this document are available:Photo images[em01007t3.jpg] [em01007t1.jpg] [em01007f1.jpg] [em01007t2.jpg] |
| |||||||||
{kind=link}
{kind=link}
{kind=link}