 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Entomotropica antes/formerly Boletín de Entomología Venezolana , Vol. 19, No. 1, April ,2004, pp.7-14 Efecto de lambdacihalotrina y metomil sobre la biología de Spodoptera frugiperda (Smith) (Lepidoptera: Noctuidae)Franklin Morillo1, Armando Notz2 1INIA.
Estación Experimental del Estado Miranda. Zona postal 1246.
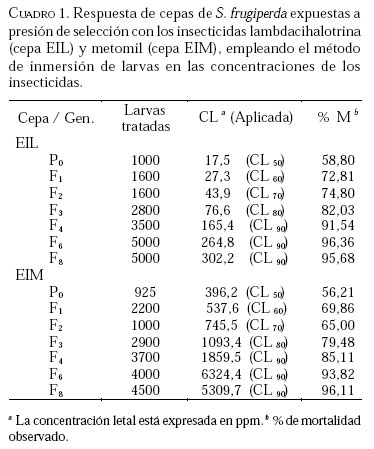
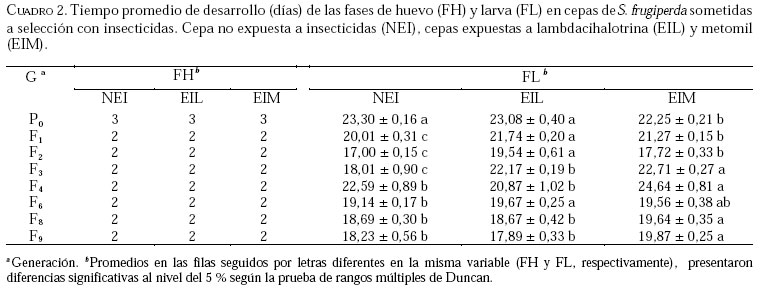
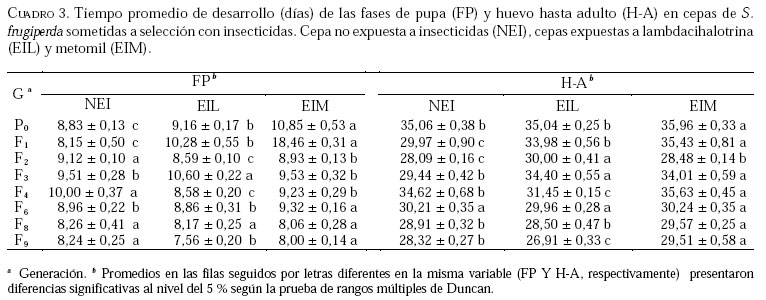
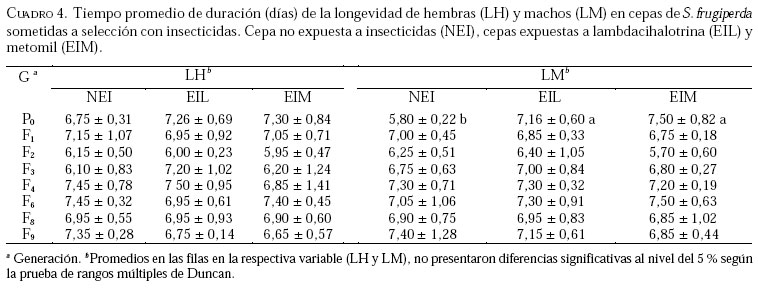
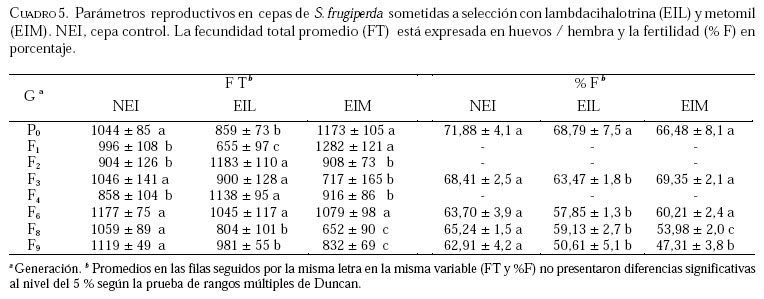
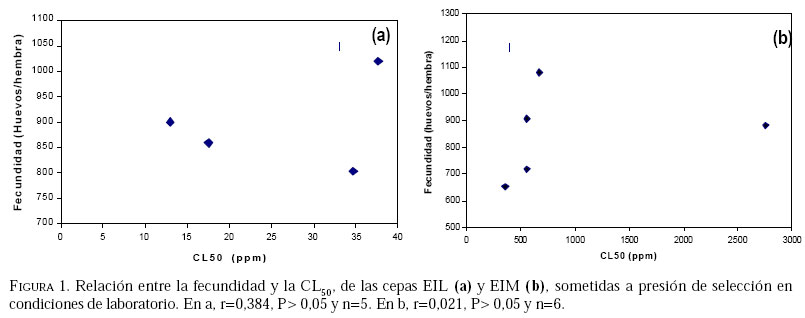
Caucagua, Edo. Miranda. Venezuela. E-mail: famorillo@inia.gov.ve, Recibido: 04-xi-2002 Code Number: em04001 Resumen MORILLO F, NOTZ A. 2004. Efecto de lambdacihalotrina y metomil sobre la biología de Spodoptera frugiperda (Smith) (Lepidoptera: Noctuidae). Entomotropica 19(1):7-14. El presente trabajo se realizó con el objeto de investigar el efecto de los insecticidas lambdacihalotrina y metomil sobre el ciclo biológico de cepas de Spodoptera frugiperda (Smith) expuestas a presión de selección. Dos cepas de S. frugiperda fueron expuestas simultáneamente a los insecticidas mencionados durante nueve generaciones. Adicionalmente, se mantuvo una cepa control no sometida a presión de selección. Se llevaron registros del tiempo de duración de las fases de huevo, larva, pupa y adulto, en cada cepa, así como la fecundidad y fertilidad. Las fases de larva y pupa mostraron mayor duración en la cepa expuesta a lambdacihalotrina (EIL) y en la cepa expuesta a metomil (EIM) con relación a la cepa control, desde la primera hasta la última generación. La longevidad de machos y hembras sólo mostró diferencias en algunas generaciones en las cepas sometidas a presión de selección. El ciclo total del insecto tendió a durar más en las cepas EIL y EIM en comparación con la cepa control. La fertilidad disminuyó a 50,61 y 47,31 % en la última generación evaluada, en las cepas EIL y EIM, respectivamente. Los resultados indican que las diferencias en la duración de algunas de las fases del insecto, representan un deterioro reproductivo en compensación de la sobrevivencia ante el proceso de presión de selección con los insecticidas lambdacihalotrina y metomil. Palabras clave adicionales: Ciclo biológico, fases de larva y pupa, gusano cogollero del maíz, insecticidas, presión de selección. Abstract MORILLO F, NOTZ A. 2004. 2004. Effect of lambdacyhalotrin and methomyl on the biology of Spodoptera frugiperda (Smith) (Lepidoptera: Noctuidae). Entomotropica 19(1):7-14. Research was conducted to evaluate the effect of the insecticides lambdacyhalotrin and methomyl on the biology of Spodoptera frugiperda (Smith) strains, exposed to selection pressure. Two strains of S. frugiperda were exposed simultaneously to the insecticides during nine generations. Additionally, we kept control strain not exposed to selection pressure. Development time of the egg, larval, pupal and adult stages in each strain, as well as the fecundity and fertility were recorded. The duration of the larval and pupal stages was significantly longer in the lambdacyhalotrin-selected strain (EIL) and the methomyl-selected strain (EIM) compared to the control strain, from the first to the last generation. The longevity of males and females only showed differences in some generations in the strains exposed to insecticides. The developmental period from egg to adult was longer in the EIL and EIM strains than in the control. The fertility diminished to 50.61 and 47.31% in the last generation, in the EIL and EIM strains, respectively. Results indicated that the differences in the duration of some of the insect phases represent a reproductive deterioration in compensation of the survival to the process of selection pressure with the insecticides lambdacyhalotrin and methomyl. Additional key words: Fall armyworm, insecticide, life history, larval and pupal stages, selection pressure. Introducción La resistencia a insecticidas en muchas especies de insectos, está frecuentemente relacionada con variaciones o cambios biológicos de comportamiento como el vigor, tiempo de desarrollo de las fases inmaduras y alteraciones del potencial reproductivo (Campanhola et al. 1991, Saume 1992). Tales modificaciones han sido observadas en especies de insectos sometidas a presión de selección con compuestos químicos. Sin embargo, estos efectos no han sido siempre consistentes, por lo que en algunos casos se presentan correlacionados positiva o negativamente con la resistencia. Algunos estudios han señalado alteraciones en las características biológicas del gusano cogollero del maíz (S. frugiperda) y otras especies relacionadas, cuando se han expuesto a la acción de insecticidas. Se ha demostrado que concentraciones subletales de diferentes compuestos xenobióticos inhiben el desarrollo en S. frugiperda con significativas variaciones de un compuesto a otro (Ross y Brown 1982). También se ha encontrado que el peso y desarrollo de larvas de S. frugiperda sometidas a aplicaciones tópicas con metomil son significativamente afectados (Javid y All 1984). En Venezuela, se ha determinado que en poblaciones de campo y laboratorio de S. frugiperda bajo presión de selección, es posible inducir resistencia a metomil (Dávila 1986; Parra 1986; Morillo y Notz 2001). Otras investigaciones han señalado ligeras alteraciones en la fecundidad y fertilidad propia de la especie, cuando es expuesta al mismo insecticida mediante pruebas de presión de selección (Anson 1987; Herrero 1987). Adicionalmente en estos trabajos se hace énfasis en la necesidad de contribuir a los estudios de resistencia a metomil y otros compuestos usados en forma inadecuada en el control de esta especie, a objeto de implementar medidas más racionales en el manejo de los insectos plaga y su respectiva resistencia. Entre otras especies afectadas se puede mencionar Spodoptera exigua, insecto en el que se demostró que el tiempo de desarrollo de larvas fue menor en la cepa susceptible en comparación a la resistente (Brewer y Trumble 1991). Además, la fecundidad fue considerablemente mayor en la primera cepa mencionada. En Heliothis virescens se reportó una tendencia similar a la anterior. Adicionalmente, se señaló que la fecundidad de hembras de la cepa susceptible fue significativamente superior a la de hembras pertenecientes a la cepa resistente (Campanhola et al. 1991). En Phthorimaea operculella (Lepidoptera: Gelechiidae) se señaló una tendencia a prolongar el ciclo de vida en cepas sometidas a selección en generaciones sucesivas (García 1989). Cepas de Spodoptera litura sometidas a dosis subletales de cihalotrina y fluvalinato, mostraron una tendencia a desarrollarse en mayor tiempo, en las fases de larva y pupa, que la cepa control (Abro et al. 1997). Sin embargo, cepas de Platynota idaeusalis (Lepidoptera: Tortricidae) expuestas a los insecticidas azinfosmetil y diflubenzuron no mostraron diferencias en el desarrollo larval, pupal ni reproductivo (Biddinger y Hull 1999). Por otro lado, la fecundidad y la viabilidad de huevos de cepas de Diatraea saccharalis (F.) (Lepidoptera: Crambidae) fue considerablemente afectada a lo largo de varias generaciones de selección, hasta el punto de perderse las cepas por la falta de oviposición (Rodríguez et al. 2001). En M. domestica, se ha señalado una reducción de la fecundidad y un período de desarrollo más largo en cepas resistentes a insecticidas (Roush y Plapp 1982), en este mismo insecto también se ha reportado una considerable reducción de la fecundidad y fertilidad en 25 generaciones de selección con el piretroide betacipermetrina (Pap y Tóth 1995). En una población de Aedes aegypti (L.) (Diptera: Culicidae), sometida a presión de selección con cipermetrina, durante cinco generaciones sucesivas, no se detectó anomalías congénitas en los sobrevivientes, ni sobre el desarrollo, comportamiento y fertilidad de los mismos (Tovar 1989). El conocimiento de la influencia de este fenómeno sobre el potencial biótico en una especie determinada, puede ser un gran aliado en el desarrollo de programas de manejo de la resistencia en las especies problema. Este trabajo tiene como propósito investigar el efecto de los insecticidas lambdacihalotrina y metomil sobre el ciclo biológico de cepas de S. frugiperda expuestas a presión de selección con los mencionados insecticidas durante varias generaciones, y establecer la correlación existente entre los valores de CL50 y la fecundidad de hembras de la especie en estudio, en las dos cepas seleccionadas. Materiales y Métodos El estudio se realizó en el insectario del Instituto de Zoología Agrícola de la Facultad de Agronomía de la Universidad Central de Venezuela, en condiciones ambientales, con promedios de temperatura y humedad relativa de 25,18 ± 5,96 °C y 75 ± 10% respectivamente, y un fotoperíodo de 12:12 horas (D:N). Los insecticidas empleados fueron el piretroide lambdacihalotrina [ciano-3-fenoxibencil-3-(2-cloro3,3,3-trifluoro-1-propanil)-2,2-dimetil-ciclopropano carboxilato] 5% C E (KARATE ®), y el carbamato metomil [S-metil-N-(metilcarbamoiloxi)tiocetimidato] 25% C E (LANNATE L ®). Cepas de S. frugiperdaevaluadas. Las cepas del gusano cogollero del maíz, utilizadas en el presente trabajo fueron las descritas en Morillo y Notz (2001). Para los bioensayos se utilizaron larvas del tercer instar de S. frugiperda, obtenidas mediante el establecimiento de una población de laboratorio, con larvas colectadas en campos de maíz en el asentamiento campesino La Cuarta, Municipio Libertador, Edo. Aragua. Las larvas fueron alimentadas y mantenidas según metodología de García (1982), los adultos fueron alimentados con una solución azucarada al 25%. Una vez multiplicada y estabilizada la cepa, se separaron tres grupos: el primero fue sometido a presión de selección con lambdacihalotrina (cepa EIL), el segundo fue tratado con el insecticida metomil (cepa EIM) y el tercero constituyó la cepa control no expuesta a insecticidas (NEI). Características biológicas registradas. Se registró la duración de todas las fases de desarrollo en nueve progenies. Las larvas sobrevivientes a las 24 horas después de aplicada cada presión de selección (Morillo y Notz 2001) fueron individualizadas y alimentadas con dieta artificial según metodología de García (1982). Las observaciones fueron realizadas diariamente utilizando una lupa binocular estereoscópica con aumento de 20x, hasta la fase de pupa. De los adultos recién emergidos en cada generación, se formaron 10 parejas de macho y hembra, cada una separada en envases de 600 cc de capacidad tapados con papel absorbente; estas parejas se alimentaron con una solución azucarada al 25% cada dos días. Al iniciarse la oviposición, la postura de cada hembra en el papel absorbente fue colectada e identificada diariamente; los contajes de huevos totales por cada hembra en todas las generaciones fueron realizados a través del peso directo de la masa de huevos alrededor de un área de 6,0 cm2 en el papel. La fertilidad se evaluó solamente en las generaciones P0, F3, F6, F8 y F9. A los datos correspondientes a duración de la fase de huevo, se les efectuó un análisis no paramétrico de una sola vía (prueba de Kruscal-Wallis), el resto de la información fue sometida al análisis de varianzas (ANAVAR) y las medias separadas de acuerdo a la prueba de rangos múltiples de Duncan a un nivel de confiabilidad del 5%. Entre las variables referidas a la fecundidad y la CL50, se computó el Coeficiente de Correlación de Pearson (r). Los análisis se realizaron con el programa estadístico computarizado SAS (SAS Institute 1985). Resultados y Discusión En el Cuadro 1, se muestra que las concentraciones subletales y letales para lambdacihalotrina y metomil aplicadas sobre S. frugiperda a lo largo del experimento en la presión de selección, variaron en la amplitud 17,5 a 302,2 ppm y 396,2 a 5309,7 ppm, respectivamente, cifras que reflejan aumentos progresivos de la toxicidad de ambos compuestos plaguicidas a medida que avanza la presión de selección. Al referirnos a la mortalidad observada, podemos apreciar que los porcentajes oscilaron entre 58,80 a 95,68% para lambdacihalotrina y entre 56,21 a 96,11% para metomil, tales valores fueron superiores a los porcentajes de mortalidad esperados en casi todas las generaciones evaluadas en ambos insecticidas. En Morillo y Notz (2001) se presenta una discusión más amplia de este aspecto. El efecto de los insecticidas seleccionados sobre la fase de huevo y larva de cepas de S. frugiperda se muestra en el Cuadro 2. El tiempo promedio de la duración de la fase de huevo no presenta diferencias significativas entre las cepas en cada generación; en P0 los huevos tardan tres días en eclosionar en cada cepa. En el resto de las generaciones sólo tardan dos días. El tiempo de desarrollo de la fase de larva en P0 fue igual entre las cepas NEI y EIL (con valores de 23,30 y 23,08 días respectivamente) y diferente a la cepa EIM (con 22,25 días); en F1 y F2 la cepa EIL fue significativamente diferente del resto de las cepas y a su vez la cepa EIM difirió de la NEI. La fase de la larva tendió a durar más tiempo en las cepas seleccionadas con productos químicos. En F3 el desarrollo larval es significativamente diferente entre las cepas, los valores extremos son 22,71 y 18,01 días en las cepas EIM y NEI, respectivamente; en F4, F8 y F9 la cepa EIM difiere significativamente del resto de las cepas, además presentó las cifras más altas de duración de la mencionada fase; en F6 la cepa EIL fue estadísticamente diferente de la cepa NEI, la EIM formó un grupo intermedio. Estos datos demuestran que en la mayoría de las generaciones la fase larval fue más tardía en las cepas seleccionadas. En el Cuadro 3 se presenta el tiempo de desarrollo promedio de la fase de pupa y el ciclo total (huevo hasta adulto). En la generación P0 las tres cepas fueron significativamente diferentes entre sí, siendo los valores extremos 10,85 y 8,83 días en las cepas EIM y NEI respectivamente; en F1 se presentó una situación similar a lo ocurrido en P0, la cepa EIM (12,46 días) y la cepa NEI (8,15 días) formaron grupos extremos, en F2 y F4 la cepa NEI presentó diferencias estadísticamente significativas respecto a las otras dos cepas, además, representó las cifras de mayor duración de la pupa; en F3 la cepa EIL fue diferente a las cepas EIM y NEI, éstas formaron un grupo igual entre sí, la fase varió de 10,60 días en EIL a 9,51 días en NEI; en F6 la duración de la pupa en la cepa EIM difirió significativamente de las otras dos cepas, las cuales formaron un grupo igual entre sí, el promedio varió de 9,32 a 8,86 días en las cepas EIM y EIL, respectivamente; en las generaciones F8 y F9 no se presentaron diferencias significativas entre las cepas para la mencionada fase. Según estos resultados, en la la mayoría de las generaciones las cepas seleccionadas presentaron la mayor duración de la pupa en comparación a la cepa control. El tiempo promedio de duración del ciclo (fase de huevo hasta adulto) en la cepa EIM fue estadísticamente diferente a las cepas EIL y NEI en la P0 (Cuadro 3). Además, estas últimas cepas fueron iguales entre sí, los valores del ciclo variaron de 35,96 días en EIM a 35,04 días en EIL y 35,06 en NEI. Una situación similar a lo mencionado anteriormente ocurre en F8, el ciclo donde varía de 29,57 a 28,50 en EIM y EIL, respectivamente; en F1 la fase del insecto difirió significativamente entre las cepas. El promedio de desarrollo osciló entre 35,43 días en EIM y 29,97 días en NEI. Èstos valores formaron los grupos extremos. En F2 se presentó una situación semejante a la anterior, sólo que los valores extremos los formaron la cepa EIL con 30,0 días y la cepa NEI con 28,09 días; en F3 las cepas EIL y EIM fueron iguales entre sí y diferentes de la cepa NEI, ésta con 29,44 días de duración representó el menor período del insecto en esta fase. En F4 existieron diferencias significativas entre las cepas; la cepa NEI formó un grupo intermedio (34,62 días) y la cepa EIL mantuvo la menor duración (31,45 días), la F9 presentó una tendencia similar a F4, sólo que la duración de los períodos en cuanto a valor absoluto fueron diferentes; en F6 no se observaron diferencias estadísticamente significativas entre las cepas. Los resultados en las fases de pupa y ciclo completo de S. frugiperda mostraron una tendencia a ser más tardíos en las cepas seleccionadas en relación con la cepa control, especialmente en las dos primeras generaciones. Adicionalmente, se indica que este efecto fue mayor cuando la cepa fue expuesta al insecticida metomil en la mayoría de las generaciones. Los resultados indican que la longevidad promedio de los adultos es poco afectada por los insecticidas (Cuadro 4). En P0 la longevidad de machos en las cepas EIM y EIL fue igual entre sí estadísticamente y diferente de la cepa NEI, los promedios son 7,50; 7,16 y 5,80 días respectivamente. En el resto de las generaciones no se observaron diferencias estadísticamente significativas de la longevidad de ambos sexos entre las cepas. La fecundidad y la fertilidad deS. frugiperda también se vieron afectadas por los insecticidas bajo estudio. La fecundidad total promedio (Cuadro 5) en P0 fue igual estadísticamente entre las cepas EIM y NEI, mientras que en la cepa EIL difirió de las anteriores, los promedios variaron entre 1 173 y 859 huevos/hembra en EIM y EIL, respectivamente. Tales cepas formaron los grupos extremos; en F1 las tres cepas difirieron entre sí en forma significativa, los valores extremos están representados en EIM (1282 huevos/hembra) y EIL (655 huevos/hembra); en F2 la cepa EIL fue significativamente diferente a EIM y NEI, éstas fueron iguales entre sí. EIL con 1 183 huevos/hembra fue el grupo extremo de mayor fecundidad; en F3 y en F8 la fecundidad en la cepa EIM fue significativamente diferente a las cepas EIL y NEI, en las dos generaciones el grupo extremo de menor fecundidad lo representó la cepa EIM con 717 y 652 huevos/hembra, respectivamente. En F4 ocurrió una tendencia semejante a lo observado en F2, EIL contiene el grupo extremo de mayor fecundidad (1 138 huevos/hembra). En la generación F6 no se detectaron diferencias significativas entre las cepas. En la generación F9 las cepas EIL y EIM fueron significativamente diferentes a la NEI. Con relación a la fertilidad se observa una disminución de la misma en comparación a la cepa NEI, con valores de 50,61 y 47,31 de huevos fértiles en EIL y EIM, respectivamente, en la última generación evaluada. Según la Figura 1 se observa que, estadísticamente, no existe correlación entre la fecundidad y las CL50 en las generaciones evaluadas en las cepas sometidas a presión de selección EIL y EIM, con r= 0,384, P>0,05 y r= 0,021, P>0,05, respectivamente. Las investigaciones realizadas enS. frugiperda ratifican el aumento del tiempo de desarrollo de cada una de las fases cuando el insecto es expuesto a concentraciones de insecticida. También se ha señalado que existen variaciones significativas de un compuesto a otro (Ross y Brown 1982, Dávila 1986, Herrero 1987). En este trabajo se demostró la disminución de la fertilidad del insecto bajo estudio, de generación a generación y se detectaron diferencias significativas en la fecundidad total al seleccionar el insecto con concentraciones de los xenobióticos utilizados. Anson (1987) señaló una tendencia de las características biológicas similar a lo obtenido en el presente trabajo, en una población de laboratorio de S. frugiperda sometida a presión de selección con metomil. Estos resultados sugieren que existe un efecto detrimental de las características biológicas de S. frugiperda, especialmente la reproductiva, cuando el insecto es sometido a presión de selección con metomil. En el caso de lambdacihalotrina, también ocurre un descenso de la fertilidad a pesar de permanecer estancada la tasa de desarrollo de resistencia. Adicionalmente, en ningún caso se observó correlación entre la fecundidad (característica reproductiva de importancia) con los valores de las CL50 registrados en Morillo y Notz (2001). En estudios similares realizados en S. exigua, Brewer y Trumble (1991) demostraron que el tiempo de desarrollo de larvas fue menor en la cepa susceptible en comparación a la resistente. EnH. virescensse reportó una tendencia similar a lo anterior, además, se señaló que la fecundidad de hembras de la cepa susceptible, fue significativamente superior a la de hembras pertenecientes a la cepa resistente (Campanhola et al. 1991). En M. domestica, también se ha demostrado una reducción de la fecundidad y un período de desarrollo más largo en cepas resistentes a insecticidas (Roush y Plapp 1982). Desde el punto de vista agronómico estas restricciones biológicas son importantes en el manejo de la resistencia. Por ejemplo se ha señalado que la inmigración de insectos susceptibles es la causa principal de la declinación anual de la resistencia en Heliothis armigera (Daly y Fitt 1990). Otros estudios han indicado que el bajo éxito reproductivo de insectos resistentes es una consecuencia de la disminución de la sensibilidad del sitio objetivo en muchas especies. Además este tipo de resistencia en H. virescens es recesiva para la susceptibilidad, por lo que la deficiencia reproductiva asociada a este tipo de herencia, en combinación con las diferencias de fecundidad, podrían combinarse para colocar a insectos resistentes en una gran desventaja en comparación a los insectos susceptibles (Campanhola et al. 1991). Sin embargo, es importante conocer el tipo de herencia ligado a cruces entre individuos resistentes y susceptibles en la mayoría de las especies de insectos de interés agrícola, ya que se crearía un gran problema al coexistir genes susceptibles con alta fecundidad y genes resistentes en los mismos individuos. Se puede concluir que no existe efecto alguno sobre el tiempo de duración de la fase de huevo de S. frugiperda con los insecticidas empleados. El efecto de los insecticidas sobre las fases de larva y pupa se expresa a partir de la F1 en las cepas seleccionadas, causando demoras en el tiempo de desarrollo de las mencionadas fases. La longevidad de machos y hembras no se ve afectada en algunas generaciones en las cepas seleccionadas. El ciclo total del insecto tiende a durar más en las cepas seleccionadas en comparación con la cepa control. En las cepas seleccionadas la disminución de la fecundidad promedio y la fertilidad fue considerable al igual que el número de generaciones afectadas, por lo que es evidente el efecto detrimental de los insecticidas sobre algunos parámetros biológicos de S. frugiperda. La fecundidad y las CL50 estimadas fueron independientes en el proceso selectivo. Agradecimientos Los autores desean expresar su más sincero agradecimiento a Nilda Yaneth Pérez, Franklin Rojas, Oscar Rivero, Ricardina Colmenares, Darjaniva Molina, por su gran colaboración en el desarrollo de este trabajo. A FUNDACITE-Aragua, por el apoyo financiero a través del proyecto DLAG-0044 y a Zéneca de Venezuela, S. A., por el suministro de algunos productos químicos empleados en los bioensayos. Referencias
© 2004- Sociedad Venezolana de Entomología The following images related to this document are available:Photo images[em04001t3.jpg] [em04001t2.jpg] [em04001t5.jpg] [em04001t4.jpg] [em04001f1.jpg] [em04001t1.jpg] |
| |||||||||
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}