 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Entomotropica , Vol. 19, No. 2, Aug ,2004, pp.91-100 Distribuição espacial de Oebalus poecilus (Dallas, 1851) (Hemiptera: Pentatomidae) durante a hibernaçãoRégis S. S. Santos1, Luiza R. Redaelli2, Lúcia M. G. Diefenbach3, Helena P. Romanowski4, Honório F. Prando5, Rita C. Antochevis2 1Depto. Estudos Agrários, UNIJUI – Rua do Comércio
3000, CEP 98700-000, Ijuí, RS.
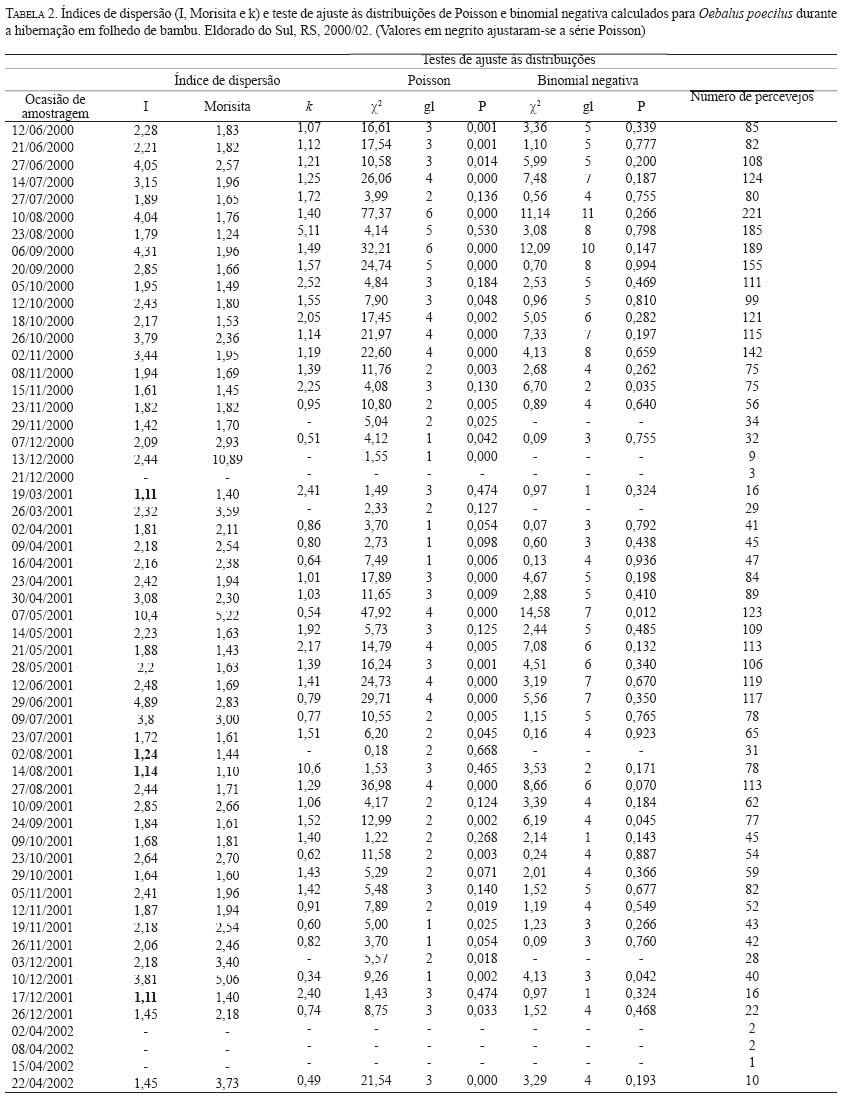
Code Number: em04014 Abstract SANTOS RSS, REDAELLI LR, DIEFENBACH LMG, ROMANOWSKI HP, PRANDO HF, ANTOCHEVIS RC. 2004. Spatial distribution of Oebalus poecilus (Dallas, 1851) (Hemiptera: Pentatomidae) in hibernation. Entomotropica 19(2):91-100. Oebalus poecilus is considered one of the most important pests in rice culture in Brazil. In this paper, the spatial distribution of this insect during its hibernation in bamboo foliage litter was studied. For the number of insects/sample unit, the adjustment of observed frequencies to the expected ones was tested, according to the Poisson, negative and positive binomial distributions; the Morisita and dispersion I indexes, the k parameter of negative binomial distribution and b of the Taylor power law were also obtained. The sample variance was higher than the average in 92.3% of the sampling occasions, with Morisita and I index statistically higher than the unit. From the occasions when the adjustment of obtained frequencies to the negative binomial distribution was tested, we could verify that only in four occasions there was no adjustment for this series. These results evidence that the pest is distributed in an aggregated mode at the hibernation site. Additional key words: Diapause, Insecta, over wintering site, small rice stink bug. Resumo SANTOS RSS, REDAELLI LR, DIEFENBACH LMG, ROMANOWSKI HP, PRANDO HF, ANTOCHEVIS RC. 2004. Distribuição especial de Oebalus poecilus (Dallas, 1851) (Hemiptera: Pentatomidae) durante a hibernação. Entomotropica 19(2):91-100. Oebalus poecilus é considerada uma das mais importantes pragas na orizicultura brasileira. Neste trabalho estudou-se a distribuição espacial deste inseto durante sua hibernação em folhedo de bambu. Para o número de insetos/unidade de amostra foi testado o ajuste das freqüências observadas às esperadas, de acordo com as distribuições Poisson, binomial negativa e positiva, também foram obtidos os índices de dispersão I e de Morisita, o parâmetro k da distribuição binomial negativa e b da lei de potência de Taylor. A variância amostral foi superior a média em 92,3% das ocasiões de amostragem, com índice I e de Morisita superiores estatisticamente a unidade. O coeficiente b de Taylor mostrou-se superior estatisticamente a unidade. Das ocasiões em que se testou o ajuste das freqüências obtidas à distribuição binomial negativa, constatouse que em apenas quatro ocasiões não houve ajuste para esta série. Esses resultados evidenciam que a praga se distribui de forma agregada no sítio de hibernação. Palavras chave adicionais: Diapausa, Insecta, percevejo-do-grão, sítio de hibernação. Introdução Entre os insetos que utilizam grãos de arroz como alimento encontra-se Oebalus poecilus (Dallas, 1851), conhecido como percevejo-do-grão. Este inseto tem ocorrência freqüente e indesejável nos arrozais, pois seus danos afetam a qualidade e a quantidade dos grãos (Santos et al. 2002), sendo apontado como importante praga em cultivos de arroz (Ferreira 1998; Gallo et al. 2002). No Rio Grande do Sul, o percevejo-do-grão tem como estratégia de sobrevivência durante o inverno a diapausa, permanecendo, neste período, em refúgios como o folhedo de bambu (Santos et al. 2002). Segundo Tauber e Tauber (1973), o entendimento das estratégias de vida dos insetos é de grande valor teórico e prático para os sistemas de manejo de pragas. Neste sentido, é necessário conhecer a biologia do inseto e sua dinâmica populacional para se elaborar um programa eficiente de manejo integrado. Assim, a elucidação de aspectos bioecológicos de O. poecilus, durante a hibernação, pode subsidiar o manejo integrado desta praga. Um dos aspectos que merece atenção é a distribuição espacial da população, a qual tem sido definida como a posição que os indivíduos ocupam, uns em relação aos outros, no habitat, em determinado momento (Southwood 1978; Ricklefs 1996). O padrão de distribuição espacial influencia a dinâmica das populações, podendo ocorrer diferenças no impacto de diversos agentes em função das variações espaciais entre os indivíduos (Heads e Lawton 1983; Hassel 1987). São três os padrões básicos de distribuição espacial das populações: agregado (ou contagioso); randômico (ou Poisson) e o regular (ou uniforme) (Bliss 1971; Elliott 1983; Ricklefs 1996). Para identificar o padrão biológico de distribuição dos organismos em suas populações, vários modelos matemáticos de distribuição de freqüência e índices de dispersão têm sido utilizados. A função Poisson tende a descrever distribuições aleatórias, enquanto a série binomial negativa a agregação (Elliott 1983; Krebs 1989). É fato bem conhecido que o padrão de distribuição ao acaso em Insecta raramente ocorre na natureza. Segundo Waters (1959) a maioria dos insetos não se distribui ao acaso no habitat, tendo em vista que poucos ambientes são homogêneos e que os fatores etológicos que determinam a sua distribuição são específicos. A tendência geral é a distribuição agregada da população nos ambientais naturais (Southwood 1978). Distribuições agregadas em heterópteros não hibernantes são freqüentemente relatadas (Kainoh et al. 1980; Foster et al. 1989; Jesus et al. 2002). A agregação fora da hibernação pode ser resultante de tendências sociais dos indivíduos em formarem grupos com diferentes finalidades: segurança, acasalamento ou reprodução (Ricklefs 1996). Em cultivos de arroz, a agregação de percevejos é relacionada ao estágio de desenvolvimento dos grãos preferidos para alimentação (Kainoh et al. 1980; Foster et al. 1989). Por outro lado, durante o período de dormência a agregação pode ter origem de variações nas condições abióticas do refúgio, proteção contra predadores e mudanças nas taxas metabólicas dos insetos (Leather et al. 1993). O padrão agregado de distribuição durante a hibernação tem sido observado e referido em diversas espécies de insetos, a maioria pertencente a ordem Coleoptera tais como: Phyllotrela cruciferae (Goeze); Psylliades punctulata Melsh. e Phyllotrela striolata (F.) (Chrysomelidae) (Burgess 1981); Oryzophagus oryzae (Costa Lima, 1936) (Curculionidae) (Mielitz 1993); Phytalus sanctipauli (Blanch., 1850) (Scarabaeidae) (Diefenbach et al. 1996). Como não há estudos sobre este aspecto durante a dormência de O. poecilus, este trabalho tem como objetivo elucidar o padrão de distribuição espacial da população hibernante deste heteróptero no sítio de hibernação. Material e Métodos O estudo foi realizado entre junho de 2000 e abril de 2002 em um taquaral formado por Bambusa tuldoides Munro (Gramineae), situado no município de Eldorado do Sul (lat 30º 02'S, long 51º 23'W), RS. O agrupamento de bambu onde o estudo foi realizado possui forma de "U"com abertura voltada para o norte. A base do "U", no sentido leste-oeste possui 100 m de comprimento por 2 m de largura. Junto à base das plantas de bambu, sobre o solo, há uma camada de folhedo com espessura média, ao longo de todo o taquaral, de 6,7 cm. Neste taquaral foram considerados para o estudo os 70 m centrais da base do "U"e, de cada lado, norte e sul, demarcadas sete áreas de 10 m de comprimento por 1 m de largura, perfazendo um total de 14 áreas. Cada uma destas 14 áreas foi subdividida em subáreas de 1x1 m, ficando cada área constituída por 10 subáreas de 1 m2. Perpendicularmente ao comprimento do bambuzal, cada subárea foi subdividida em porções de 20 cm de comprimento por 1 m de largura, num total de cinco porções de 0,2 m2 em cada metro. Um sistema estratificado aleatório de amostragem foi utilizado, onde em cada uma das 14 áreas foram retiradas quatro unidades de amostra de 300 cm3 de folhedo, das subáreas e porções, previamente sorteadas, em intervalos semanais ou quinzenais, totalizando 56 unidades de amostra por ocasião de amostragem. Em laboratório, os percevejos foram, manualmente, extraídos das amostras e contados. Para análise da distribuição espacial de O. poecilus no sítio de hibernação, primeiramente organizaram-se os dados pelas freqüências (insetos/unidade de amostra). Posteriormente, calcularam-se a média (x) e a variância (s2) para cada ocasião de amostragem, os índices de dispersão I, de Morisita e o parâmetro k (Krebs 1989). O índice de dispersão I, avalia a razão entre a variância e a média da amostra (s2/x) e indica a série Poisson quando a razão é igual a um e agregada quando essa se afasta da unidade (Elliott 1983). O índice de Morisita (Id) calculado através da expressão abaixo, indica, quando igual à unidade, o ajuste a uma série Poisson, para valores menores que um a regular e à agregada para valores maiores que a unidade (Poole 1974). 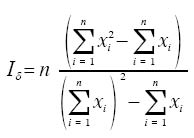
onde: n = tamanho da amostra xi= número de indivíduos na i -ésima unidade amostral O parâmetro k da distribuição binomial negativa foi estimado pelo método iterativo da "máxima verossimilhança" apresentado por Bliss e Fisher (1953): onde: n = número total de unidades de amostra ln = logaritmo natural
k= parâmetro de dispersão estimado Este parâmetro é um indicador d agregação, o qual tendendo a zero denota um distribuição agregada e ao infinito aproxima-se d acaso (Poisson) (Poole 1974). A relação entre a variância e a médi durante todo
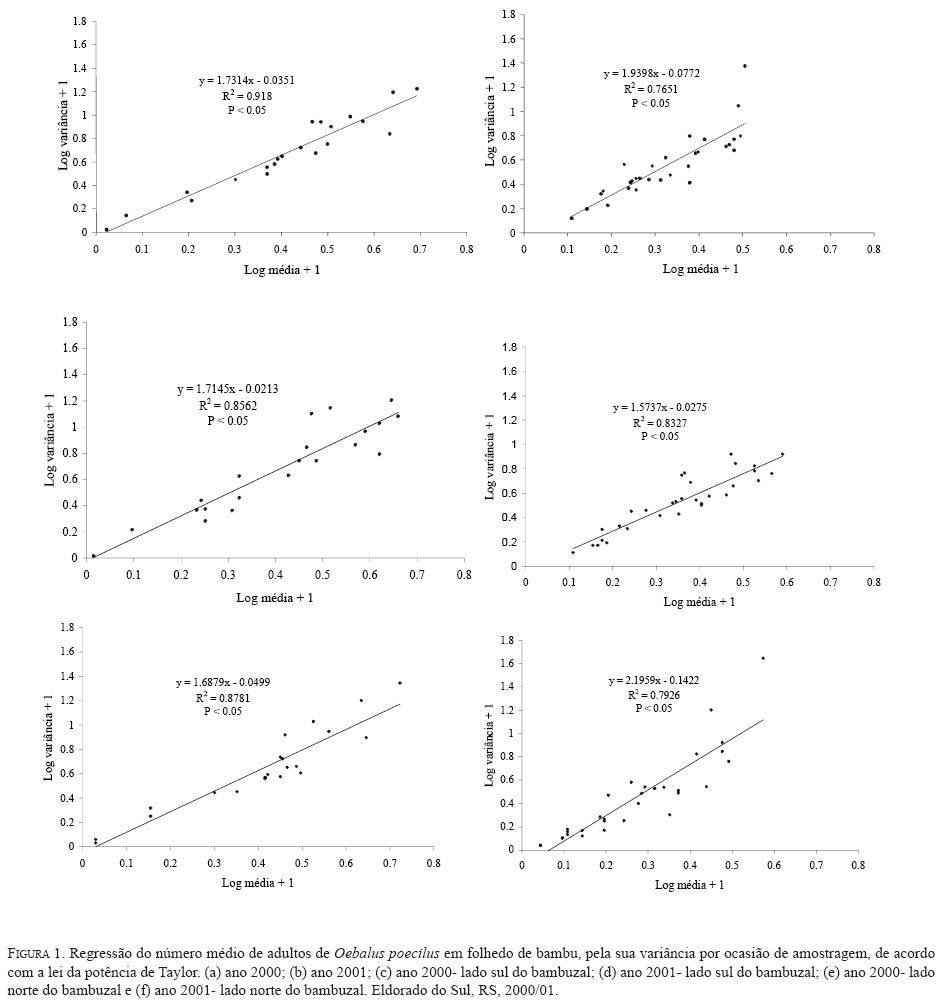
o período amostral, em cada ano, fo expressa pela lei de potência
de Taylor (log s2=log a + b log A bondade de ajuste das freqüência observadas às distribuições de Poisson, binomia negativa e positiva foi testada através da estatístic qui-quadrado (Sokal e Rohlf 1981). 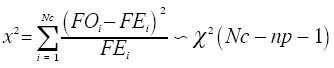
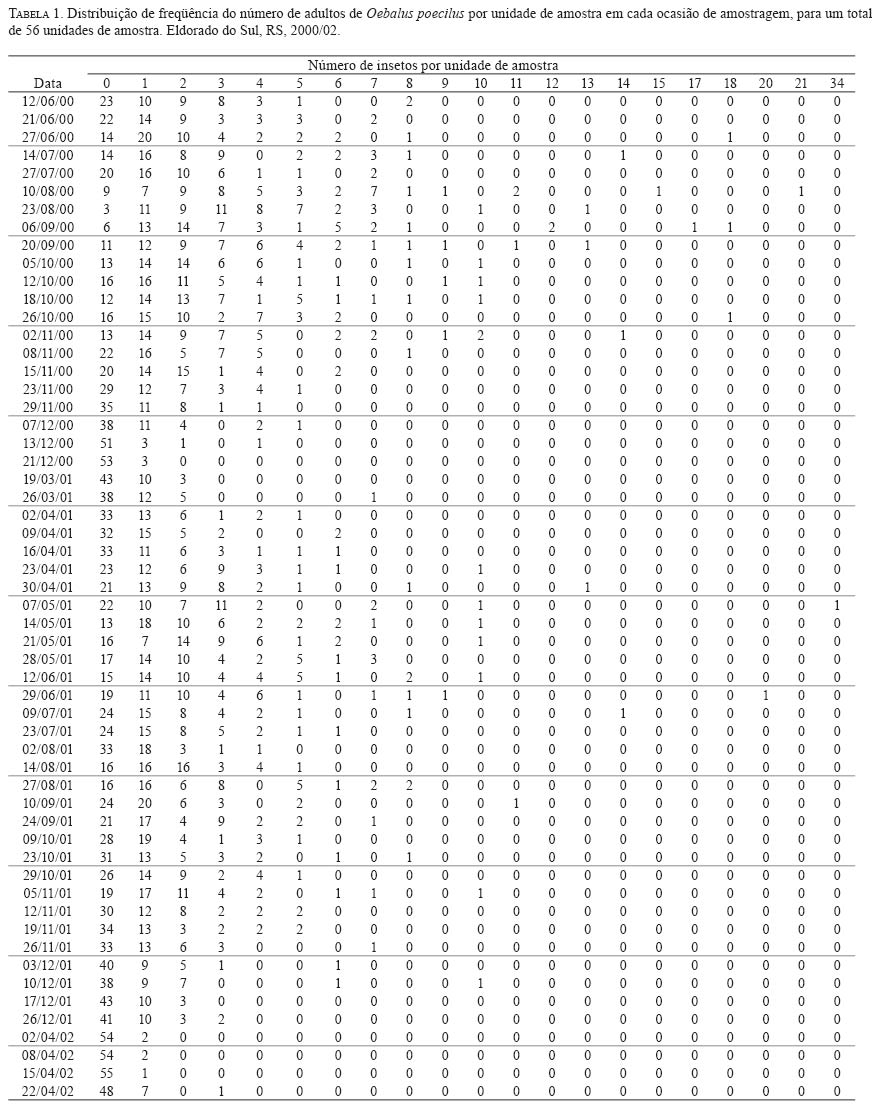
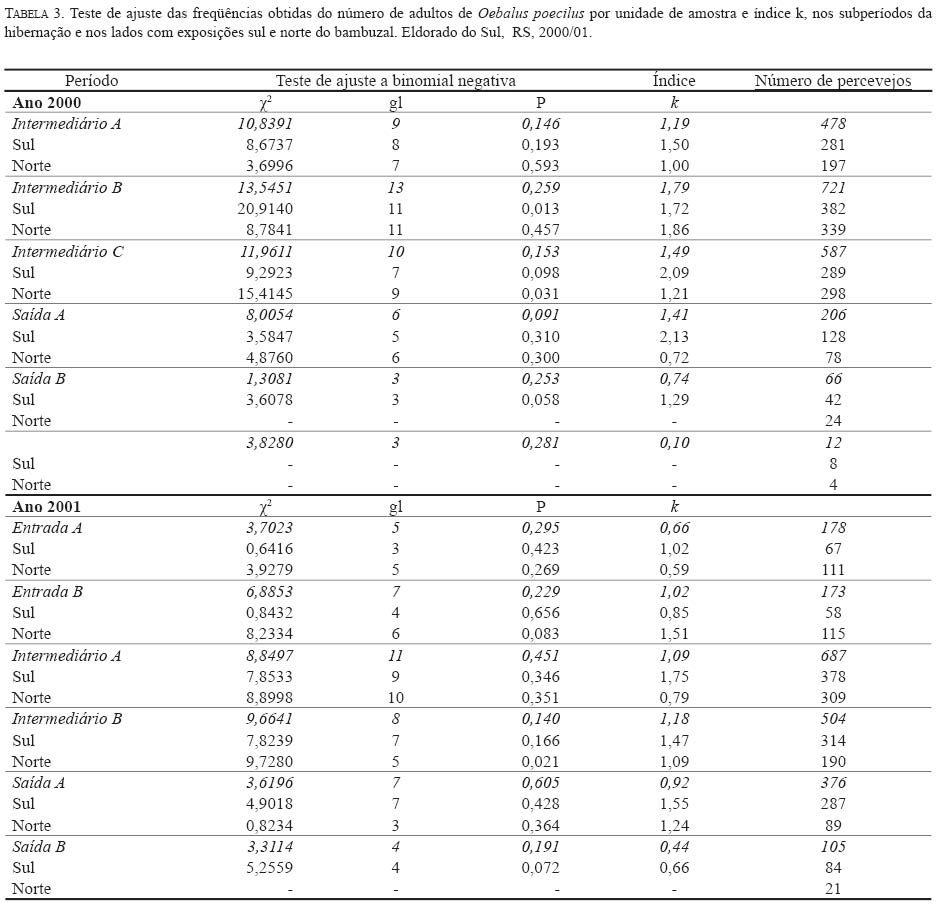
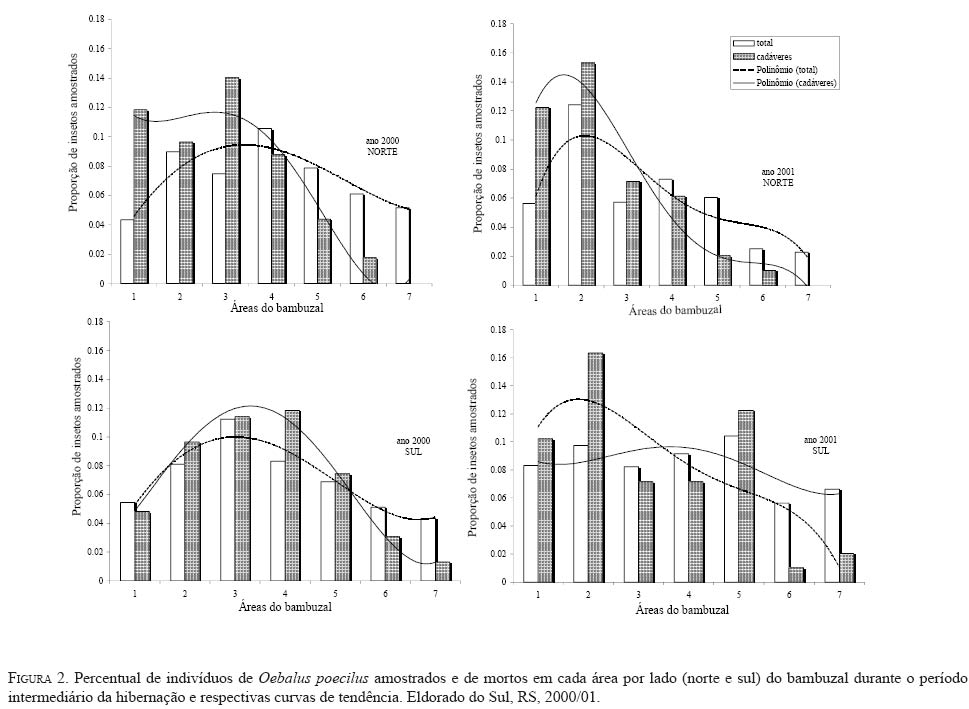
onde: Nc = número de classes da distribuição de freqüências FO = freqüência observada na i-ésima classe FE = freqüência esperada na i-ésima classe np = número de parâmetros em estudo estimados A estatística qui-quadrado também foi utilizada para verificar a significância de I. Para o parâmetro b empregou-se o teste t (Elliott 1983), e para o índice de Morisita o teste F(Poole 1974). O nível de significância adotado para todos os testes foi de 5%. A análise também foi realizada nos subperíodos da fase de hibernação quais sejam: ano 2000 -intermediário A (12/6 -27/7); intermediário B (10/8-20/9); intermediário C (05/ 10-02/11); saída A (08/11-23/11); saída B (29/11-07/12); saída C (13/12-21/12); ano 2001 -entrada A (19/3-16/ 4); entrada B (23/4-30/4); intermediário A (07/5-29/6); intermediário B (09/7-24/9); saída A (9/10-26/11); saída B (03/12-26/12). Para os cálculos foram utilizados os aplicativos Microsoft Excel, Bioestat 2.0, Krebs\Win 0.9 e SAS System. Resultados e Discussão Em todas as ocasiões de amostragem registrou-se um maior número de unidades de amostra contendo de zero a dois percevejos (Tabela 1). Unidades de amostra contendo mais do que nove indivíduos foram registradas em 22 ocasiões e apenas em oito constatou-se mais do que uma unidade de amostra contendo mais do que nove indivíduos. Em quatro ocasiões de amostragem (21/12/00; 02/04/02; 08/04/02 e 15/04/02) em função do reduzido número de indivíduos amostrados (menor do que quatro) que resultou em apenas duas classes de freqüências, não foi possível realizar o cálculo do ajuste das freqüências observadas aos modelos de distribuição e índices de dispersão utilizados. Além destas ocasiões em mais cinco (29/11/00; 13/12/00; 26/03/01; 02/08/01 e 03/12/01) o número reduzido de classes de freqüências também não permitiu o teste de ajuste à série binomial negativa. Com exceção do dia 02/ 8/01, essas ocasiões incluem-se nos períodos de entrada ou saída do sítio de hibernação quando a densidade de O. poecilusé naturalmente baixa no refúgio. Analisando as ocasiões de amostragem em que o número de indivíduos amostrados permitiu cálculo estatístico, a variância do número de percevejos por unidade de amostra, representada pelo índice I, mostrou-se estatisticamente superior à média em 48 ocasiões (92,3%; P<0,001), indicando o padrão espacial de distribuição agregado do percevejo-do-grão durante a hibernação. Este tipo de distribuição também é evidenciado pelo índice de Morisita em todas as datas de amostragem (P<0,05) (Tabela 2). Nas quatro ocasiões em que o índice I aponta para uma distribuição aleatória (19/03/01; 02/08/01; 14/08/01 e 17/12/01), verificou-se o ajuste simultâneo a Poisson e, quando o número de insetos permitiu seu cálculo, também ao modelo binomial negativo. Nestas ocasiões, excetuando-se o dia 14/08/01, um número reduzido de indivíduos (menor do que 31) foi amostrado, o que pode explicar o resultado encontrado. Segundo Southwood (1978) quando a população é muito esparsa as chances dos indivíduos ocorrerem em qualquer unidade amostral são tão pequenas que sua distribuição é efetivamente ao acaso. Turnock et al. (1987) em amostragens de serapilheira em Manitoba, Canadá, constataram em anos em que a densidade populacional média por unidade de amostra foi maior do que um, a distribuição dos crisomelídeos P. cruciferae e P. striolata foi agregada. Entretanto, em densidades menores do que um a distribuição seguiu o padrão ao acaso. Boeve and Weiss (1998) atribuem a não agregação de Shizaphis graminum (Rondani, 1852) (Hemiptera: Aphididae) ao reduzido tamanho populacional da praga em campos de trigo, que resultou em um baixo coeficiente de determinação pela lei de potência de Taylor. Em todas as ocasiões em que se efetuou teste de ajuste das freqüências obtidas aos modelos matemáticos propostos, não se evidenciou o ajuste a série binomial positiva. Segundo Southwood (1978) a disposição regular que o modelo indica é rara em populações animais. O padrão de distribuição regular é freqüentemente observado em populações artificiais tais como culturas, onde as plantas estão uniformemente dispostas, de modo que o número de indivíduos em cada unidade de área é o mesmo (Taylor 1965). Das 47 ocasiões em que se testou o ajuste das freqüências obtidas à distribuição binomial negativa, constatou-se que não houve o ajuste em apenas quatro ocasiões (15/11/00; 07/05/01; 24/09/01 e 10/12/01). Apesar de não mostrar ajuste à série binomial negativa, os valores do índice k são próximos de zero, indicando a agregação, além disso, os índices I e de Morisita confirmam o padrão agregado de O. poecilus nestas datas. Mielitz (1993) estudando a distribuição espacial de adultos de O. oryzae hibernantes, encontrou situação semelhante à anterior. A autora sugere que os pequenos valores de k, mesmo não ocorrendo ajuste ao modelo binomial negativo, indicam o padrão agregado do inseto, e que, provavelmente, algum outro modelo de distribuição contagiosa possa descrever a agregação. Conforme Maruyama et al. (2002) o modelo de distribuição binomial negativo é muito flexível, ajustandose a uma ampla faixa de distribuições, que vão da altamente agregada até aquelas tendendo à aleatoriedade, dependendo do valor do parâmetro k. O valor de k pode variar de zero ao infinito, quando k é próximo a zero maior é a agregação da população, já valores de k superiores a oito, a variância se aproxima da média e a distribuição binomial negativa converge para Poisson (Bliss 1971; Poole 1974). Assim, os resultados aqui obtidos em relação ao ajuste à série binomial negativa, e aos índices I e de Morisita, confirmam a hipótese de que o padrão espacial de distribuição apresentado pelos adultos de O. poecilus ao longo da hibernação foi o agregado. A agregação do percevejo-do-grão também é sustentada pela lei da potência de Taylor tanto em 2000 (b=1,7314; t=6,898; gl =19; P<0,05) quanto em 2001 (b=1,9398; t=4,708; gl =29; P<0,05), com o modelo de distribuição agregado explicando 91,8% e 76,5% da variação observada em 2000 e 2001, respectivamente. (Figura 1). Quando se analisou a distribuição espacial de O. poecilus considerando-se os subperíodos da hibernação, a tendência de agregação torna-se ainda mais marcante, com ajuste a série binomial negativa em todos os subperíodos e valores de k ao redor de 1,19. (Tabela 3). O padrão agregado do percevejo-do-grão também pôde ser constatado nos lados do bambuzal com exposições norte e sul na maioria dos subperíodos (Tabela 3). Cabe ressaltar que para execução da análise por lado do bambuzal, a redução no número de unidades de amostra impediu a análise dos dados em alguns subperíodos da saída do refúgio, quando um pequeno número de insetos foi coletado. Pela lei de potencia de Taylor também se evidenciou o padrão agregado do percevejo-do-grão tanto no lado sul do bambuzal em 2000 (b=1,7145; t=4,435; gl =19; P<0,05) e 2001 (b=1,5737; t=4,382; gl =29; P<0,05) quanto no norte em 2000 (b=1,6879; t=4,767; gl =19; P<0,05) e 2001 (b=2,1959; t=5,735; gl =29; P<0,05), com o modelo explicando pelo menos 79,3% da variação observada (Figura 1). O padrão de distribuição espacial agregado registrado para o percevejo-do-grão corrobora o apontado para as espécies hibernantes P. cruciferae; P. punctulata; P. striolata (Burgess 1981); O. oryzae (Mielitz 1993) e P. sanctipauli (Diefenbach et al. 1996). A agregação durante a hibernação parece ser resultante de inúmeros fatores tais como: proteção às baixas temperaturas (Holmquist 1931); existência de uma condição abiótica preponderante para a população no local (Bullock 1967); disponibilidade limitada de refúgios apropriados (Danks 1978) ou respostas comportamentais à heterogeneidade do habitat em relação a fatores como umidade, temperatura, sombreamento e ação eólica (Mielitz 1996). Segundo Denlinger (1986) os agregados modificam, significativamente, as condições do microclima no refúgio para o inseto por reduzir a evaporação da água. Em ambientes que apresentam certos níveis de umidade a temperatura é amenizada e, conseqüentemente, as injúrias por frio ou a morte são minimizadas (Dixon et al. 1979). Segundo Holmquist (1931) e Leather et al. (1993) os refúgios de hibernação são escolhidos pelos insetos por apresentarem uma menor amplitude de variação da temperatura e de umidade relativa do ar, o que segundo Salin et al. (2000) gera um ambiente mais estável para o inseto hibernar. Entretanto, em função da espessura da serapilheira e da orientação solar e eólica do refúgio estes dois fatores podem apresentar diferenças em pontos distintos do sítio (Mielitz 1993). Assim, o padrão agregado constatado para O. poecilus pode estar relacionado a uma provável heterogeneidade do folhedo de bambu fazendo com que O. poecilus prefira determinados locais em detrimento de outros. Na Figura 2 estão apresentados os percentuais de insetos amostrados e de indivíduos mortos em cada uma das áreas de estudo durante o período intermediário da hibernação de O. poecilus. Em geral, houve uma tendência da população em permanecer nas porções mais centrais da área experimental em ambos os lados do bambuzal. Nas áreas extremas (6 e 7), em ambos os lados evidenciouse uma menor preponderância da população. Segundo Danks (1978) altas taxas de mortalidade são esperadas para os insetos que não hibernam no local mais favorável. No presente estudo o oposto foi constatado, áreas com maior concentração de insetos apresentaram os maiores percentuais de morte em ambas as exposições do bambuzal (Figura 2). Este fato pode explicar o ajuste das freqüências obtidas do número de insetos mortos por unidade de amostra, a série binomial negativa em praticamente todos os subperíodos da hibernação, com índice k em torno de 0,98, o que evidencia o padrão de distribuição agregado também dos cadáveres do percevejo-do-grão. O exame dos dados da Tabela 3 e do número de adultos de O. poecilus amostrados nos dois lados do bambuzal, nos diferentes subperíodos, permite supor que o percevejo-do- grão tenha ocupado o folhedo de bambu, preponderantemente, pelo lado norte e também tenha abandonado o refúgio primeiro por este lado. Cabe ressaltar que o lado com a exposição norte é protegido dos ventos de direção leste e sudoeste, predominantes na região, além disso, tanto no período de entrada quanto no de saída registrou-se a presença de uma vegetação espontânea abundante junto a este lado, onde constatouse, em ambos os períodos, adultos do percevejo-do-grão alimentando-se. Durante o período intermediário, nos dois anos de estudo, no lado com exposição sul registrou-se sempre um maior número de adultos do percevejo-do-grão. Este fato pode estar relacionado a alguma condição favorável do ambiente no lado sul. De fato, o lado sul é o mais exposto aos ventos predominantes na região, o que pode favorecer a manutenção de um ambiente menos úmido. Além disso, este lado era um pouco mais elevado que o norte, o que pode ter auxiliado a reduzir a umidade do solo próximo ao folhedo. Tais aspectos podem ter tido infl uência no maior número de indivíduos registrados nos períodos intermediários do processo de hibernação. Um padrão agregado de distribuição associado a uma condição de maior insolação e baixa umidade foi registrado para Pissodes sorti L. (Coleoptera: Curculionidae) e L. (Coleoptera: Curculionidae) e O. oryzae nos seus sítios de hibernação, respectivamente por Dixon et al. (1979) e Mielitz (1993). O padrão de distribuição espacial agregado, encontrado para O. poecilus durante a fase de hibernação, elucida um aspecto importante da dinâmica da população o qual terá papel importante em futuros programas de manejo que visem a redução populacional do percevejo-do-grão durante a hibernação. Agradecimentos O presente estudo teve o apoio do Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq-Brasil) e da Fundação de Amparo a Pesquisa do Estado do Rio Grande do Sul (FAPERGS). Referências
The following images related to this document are available:Photo images[em04014t1.jpg] [em04014t3.jpg] [em04014t2.jpg] [em04014f2.jpg] [em04014f1.jpg] |
| |||||||||
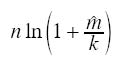
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}