 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Entomotropica antes/formerly Boletín de Entomología Venezolana, Vol. 20, No. 1, April, 2005, pp. 29-38 Caracterización de criaderos de anofelinos (Diptera: Culicidae) vectores de malaria del Alto Orinoco, Amazonas, Venezuela Yasmin Rubio-Palis1,2, Cristóbal Menare3, Andrés Quinto3, Magda Magris4, Manuel Amarista5 1Dirección
de Salud Ambiental, MSDS. Code Number: em05006 Resumen RUBIO-PALIS Y, MENARE C, QUINTO A, MAGRIS M, AMARISTA M. 2005. Caracterización de criaderos de anofelinos (Diptera: Culicidae) vectores de malaria del Alto Orinoco, Amazonas, Venezuela. Entomotrtopica 20(1): 29-38. Entre Octubre 1999 y Enero 2000 se identificaron y caracterizaron los habitats de los estadíos inmaduros de anofelinos en el área de Ocamo, municipio Alto Orinoco, Amazonas, Venezuela. Se tomaron 46 muestras de cuatro tipos de criaderos: laguna, pozo, caño y charco. Se colectaron 1 304 larvas de anofelinos de diferentes estadios, pertenecientes a: Anopheles oswaldoi, A. darlingi, A. triannulatus, A. braziliensis, A. mattogrossensis, A. punctimacula, A. forattinii y A. peryassui. La especie más abundante fue A. oswaldoi, seguida por A. triannulatus y A. darlingi. Las demás especies representaron 11% del total identificado. La densidad de larvas fue mayor en los criaderos tipo pozo (3.38 larvas/cucharón). El análisis de correspondencia canónica (ACC) realizado evidenció una fuerte estructuración espacial en relación con las especies de anofelinos y variables ambientales. Este resultado fue ratificado a posteriori por la prueba de permutación de Monte Carlo, mostrando un nivel de significación de p= 0,01. Las variables que explican la distribución espacial de A. oswaldoi, A. darlingi y A. mattogrossensis son: detritos (palitos y hojas), pH, profundidad y temperatura. A. triannulatus mostró preferencia por las lagunas con vegetación flotante y gramíneas altas. La densidad de estadios inmaduros DEL3, DEL4 y DEP-PA, es mayor en las lagunas con sombra permanente y abundantes algas. Palabras clave adicionales: Anopheles oswaldoi, A. triannulatus, A. darlingi, A. mattogrossensis, A. punctimacula, A. forattinii, A. braziliensis, A. peryassui. Abstract RUBIO-PALIS Y, MENARE C, QUINTO A, MAGRIS M, AMARISTA M. 2005. Caracterización de criaderos de anofelinos (Diptera: Culicidae) vectores de malaria del Alto Orinoco, Amazonas, Venezuela. Entomotrtopica 20(1): 29-38. Between October 1999 and January 2000, anopheline breeding habitats were identified and characterized in the Ocamo area, municipality Upper Orinoco, Amazonas state, in southern Venezuela. During the study, 46 samples were taken from 4 types of larval habitats: lagoon (37), pond (2), stream (6) and pool (1). A total of 1 304 anopheline larvae were collected belonging to 8 species: Anopheles oswaldoi, A. darlingi, A. triannulatus, A. mattogrossensis, A. punctimacula, A. forattinii, A. braziliensis and A. peryassui. The most abundant and frequent species collected was A. oswaldoi, followed by A. triannulatus and A. darlingi. The remaining species accounted for 11% of the total anophelines identified. Larvae density was higher in the ponds (3.38 larvae/dip). The analysis of canonic correspondence (ACC) showed a strong spatial structure in relation to anopheline species and environmental variables. These results were confirmed a posteriori by the Monte Carlo permutation test, p= 0,01. The variables that explained the spatial distribution of A. oswaldoi, A. darlingi and A. mattogrossensis were: detritus (twigs and leafs), pH, water depth and temperature. Anopheles triannulatus preferred lagoons with floating vegetation and tall grasses. Higher density of inmature stages (DEL3, DEL4 and DEP-PA) were associated with shaded lagoons and abundant algae. Additional key words: Anopheles oswaldoi, A. triannulatus, A. darlingi, A. mattogrossensis, A. punctimacula, A. forattinii, A. braziliensis, A. peryassui. Introducción En los últimos años se ha registrado un incremento en el número de casos de malaria en Venezuela, reportándose para el año 2004 un total de 46 244 casos, colocando al país en situación de epidemia (Reporte Epidemiológico, semana N° 52, MSDS 2004). Los estados con mayor casuística fueron Bolívar (59,6%), Amazonas (23.2%) y Sucre (10,3%), siendo Amazonas el estado con mayor transmisión al obtenerse un índice parasitario anual (IPA) de 102.2 casos por 1 000 habitantes (Reporte Epidemiológico, semana N° 52, MSDS 2004). La malaria en el municipio Alto Orinoco, ubicado en el sureste del estado Amazonas, representa una de las principales causas de morbilidad y mortalidad en las comunidades indígenas Yanomami (Informe Epidemiológico de los Ambulatorios Rurales Tipo II (ARII) de Ocamo, Mavaca y Platanal, MSDS I993-I996). Este municipio ha sido considerado como área de alto riesgo a malaria (Aché 1998). Estudios entomológicos realizados en el Alto Orinoco señalan que A. (Nyssorhynchus) darlingi Root, 1926, es el principal vector de los parásitos maláricos (Rubio-Palis et al. I997). Esta especie presenta comportamiento endofágico y exofílico, es decir, que pica dentro de las viviendas pero reposa fuera de ellas, lo cual dificulta su control con la medida tradicional de rociamiento intradomiciliario con insecticidas de acción residual. Además de esto, las viviendas de los indígenas Yanomami no presentan superficies rociables adecuadas, ya que las paredes son incompletas y fabricadas de palma, manaca y barro. La malaria es un problema de salud pública con grandes dificultades para su control en el Alto Orinoco. Es por ello que en los últimos años se han incrementado los esfuerzos por conocer y caracterizar mejor al principal vector, A. darlingi, así como otras especies que podrían representar vectores potenciales, con la finalidad de diseñar un programa de control adecuado a las características ambientales, biológicas del parásito y vector, así como a las características socioculturales de la población Yanomami (Magris et al. 1999). En este sentido es de vital importancia, intensificar la investigación sobre la biología y ecología de los anofelinos, caracterizar y ubicar sus habitats larvarios, y su posible participación en la transmisión de la malaria en estos remotos lugares. A fin de contribuir al conocimiento de la ecología de los estadíos inmaduros de los anofelinos del Alto Orinoco, se procedió a identificar y caracterizar los criaderos, así como a establecer los parámetros bióticos y abióticos que determinan la presencia o ausencia de las diferentes especies encontradas. Materiales y Métodos Área de estudio. El estudio se realizó en los alrededores de Ocamo (lat 02° 50' N, long 65° 14' W); área ubicada en el municipio Alto Orinoco, al sur-este del estado Amazonas que ocupa una extensión de 30 850 km2 (Figura 1). Debido a que no hay disponibles mapas del área a escala 1:5 000, fue necesario producir nuestro propio mapa utilizando el programa MapInfo Professional 7.0 (2001) para lo cual se registraron las coordenadas en los ríos, "shabonos" y sitios de muestreo con un GPS (Garmin GPS 12, Garmin, Olathe, KS, USA). En esta región se estima que hay unas 20000 personas de las cuales 14000 pertenecen a la etnia Yanomami (OCEI 1993). El área de Ocamo tiene aproximadamente 29,5 Km2 y se encuentra en la parte baja del Alto Orinoco, a una altitud de 116 msnm y está caracterizada por un mosaico de bosque inundado, incluyendo muchas especies de palmeras (Steyermark et al. 1995). El área de Ocamo está conformada por 11 comunidades o "shabonos" ubicados a lo largo de los ríos Orinoco (Yohope, Shashana, Lechosa y San Benito) y Ocamo (Santa Maria de los Guiacas - Ocamo, Tumba, Carlitos, Clavotheri, Kashora, Ishinopoitheri y Mario) (Figura 1). Esta región está caracterizada como selva húmeda tropical (Ewel et al. 1976), presentando una temperatura media anual de 28 °C y precipitación media anual de 3 500 – 4 000 mm (Estación Climatológica en Ocamo, MARNR 2000). La temperatura está asociada estrechamente con la distribución anual de las lluvias. Los meses con mayor precipitación y más bajas temperaturas son junio, julio y agosto, mientras que enero, febrero y marzo presentan las más altas temperaturas y menor precipitación. Eco-epidemiologicamente la zona corresponde a la clasificación de malaria de bosques bajos interiores (Rubio-Palis y Zimmerman 1997). El principal río de este municipio es el Orinoco (Figura 1), el cual nace en el cerro Delgado Chalbaud, siendo sus principales afluentes Orinoquito, Manaviche, Mavaca, Ocamo, Padamo y Cunucunuma. El río Ocamo, uno de los mayores afluentes del Orinoco, proviene de la vertiente de la Sierra Parima, aunque con matices diferentes, la mayoría de las aguas del Alto Orinoco son amarillentas. Este río se caracteriza por ser de agua clara, particularmente tendiendo a color amarillo–marrón; suspensión de sólidos y nutrientes relativamente bajos, pH casi neutro. Estas condiciones reflejan contenido de mineral de origen rocoso (ígneas, metamórficas, basamento de granito cubiertos con capa de arena) (Rejmánková et al. 1999). Cuando el agua de la inundación retrocede, se forman lagunas del río que duran hasta la próxima inundación. Durante el período de aislamiento, el nivel del agua está controlado principalmente por las precipitaciones y evapotranspiración (Hamilton y Lewis 1990). Además de las lagunas, en la selva inundada (que usualmente representa un hábitat transitorio que se forma después de las lluvias), se encuentran pequeños charcos y riachuelos que representan un hábitat potencial para las larvas. Las lagunas mantienen diversas vegetaciones acuáticas que incluyen macrófitas emergentes (graminoides, Ludwigia spp, Montrichardia arborescens), comunidades de plantas sumergidas representadas por los géneros Elodea, Mayaca, Utricularia y Cabomba, y escasamente macrofitas flotantes (Lemna y Salvinia) (Rejmánková et al. 1999). Las inundaciones en la selva proporcionan un hábitat dominado por detritus y acumulación de desechos orgánicos en la superficie del agua (Rejmánková et al. 1999). Descripción de criaderos. La descripción de los hábitats larvarios fue llevada a cabo mensualmente a lo largo de los ríos Ocamo y Orinoco (Figura 1), a partir del mes de octubre de 1999 hasta enero de 2000, correspondiendo con el período final de lluvias e inicios de la sequía. Las muestras fueron colectadas al inspeccionar cuerpos de agua alrededor de los "shabonos" y a través de la selva en criaderos previamente identificados por Rejmánková et al. (1999), así como en otros criaderos identificados durante los cuatro meses que duró el estudio. Una vez localizado el criadero se procedía a realizar la descripción del sitio de colecta tomando en cuenta la presencia y abundancia de detritus, plantas flotantes, emergentes y sumergidas, luego se procedía a realizar las mediciones de los factores abióticos temperatura, pH y profundidad. La temperatura del agua se midió con termómetro de bulbo para agua calibrado en °F, que posteriormente fue convertida en °C; el pH se midió con cintas calibradas Sigma No P-4536 con un rango de pH de 4,5 – I0,0 y la profundidad se midió con una vara marcada en metros. Colecta de mosquitos. Una vez descrito el criadero, se procedía a la colecta de larvas con el cucharón de cono truncado de 1 500 ml de capacidad. En cada sitio o punto de colecta, se realizaron 30 inmersiones del cucharón. Las larvas colectadas en cada inmersión se identificaron como pertenecientes al género Anopheles por su característica posición de reposo paralela a la superficie del agua. Para cada cucharonada se clasificaban y contaban las larvas de acuerdo al estadío (I, II, III y IV instar) y pupas. Las larvas colectadas fueron colocadas en bolsas de plástico con agua del criadero, rotuladas y transportadas al laboratorio de campo (Ambulatorio ARII Ocamo). Debido a la imposibilidad de identificar a nivel de especie los estadios larvales del I al III, se procedía a colocar estas larvas en recipientes plásticos de 300 ml de capacidad hasta alcanzar el estadío IV, cuando los especimenes eran colocados individualmente en tubos plásticos de 9 dram (9 cm alto x 2 cm diámetro) hasta completar su desarrollo en adultos, obteniéndose de esta forma para cada individuo los estadios asociados. Durante el desarrollo larvario las exuvias de larvas de IV estadio y pupas, producto de cada muda fueron preservadas en etanol al 80% para su posterior montaje permanente y su identificación a nivel de especie utilizando las claves de Navarro (1996) y Wilkerson y Sallum (1999). Los anofelinos adultos fueron montados en alfileres entomológicos # 3 y se determinó la especie con las claves para adultos de Cova García y Sutil (1977), Rubio- Palis (2000) y Wilkerson y Sallum (1999). Cada ejemplar fue debidamente identificado por especie, localidad, fecha y nombre de la persona que lo identificó. Los especimenes identificados se depositaron en la Unidad de Estudios Entomológicos del Servicio de Vigilancia Epidemiológica Sanitario Ambiental de Malariología Región XIX, Puerto Ayacucho, estado Amazonas. Se colectaron muestras de plantas acuáticas sumergidas y flotantes presentes en los criaderos tipo laguna. Las muestras fueron rotuladas con los datos del criadero y envueltas individualmente en papel periódico impregnado con etanol al 80% y colocadas en prensa hasta su traslado al Herbario del Ministerio del Ambiente en Puerto Ayacucho, para su posterior montaje y clasificación. Análisis de los datos. A fin de determinar cuales de los factores bióticos y abióticos considerados condicionan la presencia de las especies de Anopheles colectadas, se realizó un Análisis de Correspondencia Canónica (ACC) (Ter Braak I987a, b). Para probar la hipótesis nula planteada que corresponde a la ausencia de estructura canónica, se realizó la prueba de permutación de Monte Carlo (Ter Braak I987a, b). Resultados Especies de anofelinos colectadas. Durante todo el estudio se colectaron un total de 1 304 larvas de anofelinos de diferentes estadios, de estas solamente se identificaron 282 especimenes, el resto no se pudo identificar a nivel de especie, debido a que pertenecían a estadios menores al IV y no se logró que completaran el desarrollo hasta adultos. Se identificaron un total de ocho especies del género Anopheles; de éstas, cuatro pertenecientes al subgénero Nyssorhynchus: A. darlingi Root, 1926, A. oswaldoi (Peryassú, 1922), A. triannulatus (Neiva y Pinto, 1922) y A. braziliensis (Chagas, 1907) y cuatro especies al subgénero Anopheles: A. mattogrossensis Lutz y Neiva, 1911, A. punctimacula Dyar y Knab, 1906, A. peryassui Dyar y Knab, I908, y A. forattinii Wilkerson y Sallum, I999 (Cuadro 1). Cuadro 1. Número total de larvas identificadas y sus frecuencias (entre paréntesis)(*) por tipo de criadero para cada especie de anofelino. Ocamo, municipio Alto Orinoco, estado Amazonas. Octubre 1999 – Enero 2000.
(*) porcentaje de criaderos positivos a cada especie por tipo de criadero De un total de 282 larvas identificadas se destacan como especies más abundantes A. oswaldoi con 142 (50,5%) individuos, además esta especie resultó ser la más frecuente, encontrándose en 23 de las 46 muestras tomadas en los cuatro tipos de criaderos. En segundo lugar encontramos a A. triannulatus con 71 larvas identificadas (25,3%), y la tercera especie más abundante fue A. darlingi con 41 larvas (14,6%). Las demás especies colectadas fueron A. mattogrossensis (4,6%), A. punctimacula (2,8%), A. forattinii (1,8%), A. braziliensis (0,4%) y A. peryassui (0,4%) (Cuadro 1). Tipos de criaderos de anofelinos evaluados. Durante el estudio fue posible identificar y caracterizar cuatro tipos de criaderos: lagunas, caños, pozos y charca. Entre Octubre de 1999 y Enero de 2000 se tomaron un total 46 muestras distribuidas de la siguiente forma: lagunas (37), caños (6), pozo (2) y charca (1) (Fig. 1, Cuadro 2). Cuadro 2. Resultados de las colectas y densidad de anofelinos por tipo de criaderos en el área de Ocamo, municipio Alto Orinoco, estado Amazonas. Octubre 1999 - Enero 2000.
(*) Número promedio de larvas colectadas dividido entre 30 cucharones tomados en cada sitio de muestreo Laguna. Son de origen natural y en su mayoría se forman por desbordamiento y embalses de ríos y caños en depresiones naturales del terreno. Estos habitats tenían menos sombra que los demás habitats inspeccionados (pozo, caño, charca). La mayoría de estas lagunas son temporales, pero a pesar de la sequía en la época de verano (febrero, marzo), algunas de éstas mantienen el agua permitiendo así de esta manera la sobrevivencia de los anofelinos. El agua es de color marrón claro, con abundante detritus y materia orgánica, la mayoría son estancadas pero en la época lluviosa, con el aumento del nivel del río presentan corriente lenta siguiendo su curso al río. La temperatura media del agua fue de 26,5°C, profundidad variable entre 10 cm y 4 metros, y el promedio de pH del agua fue de 6,2 (Cuadro 3). Cuadro 3. Media y desviación estándar (entre paréntesis) de la variables físico-químicas medidas y bióticas (porcentaje de cobertura) estimadas en los diferentes tipos de criaderos estudiados en el área de Ocamo, municipio Alto Orinoco, estado Amazonas. Octubre 1999 - Enero 2000.
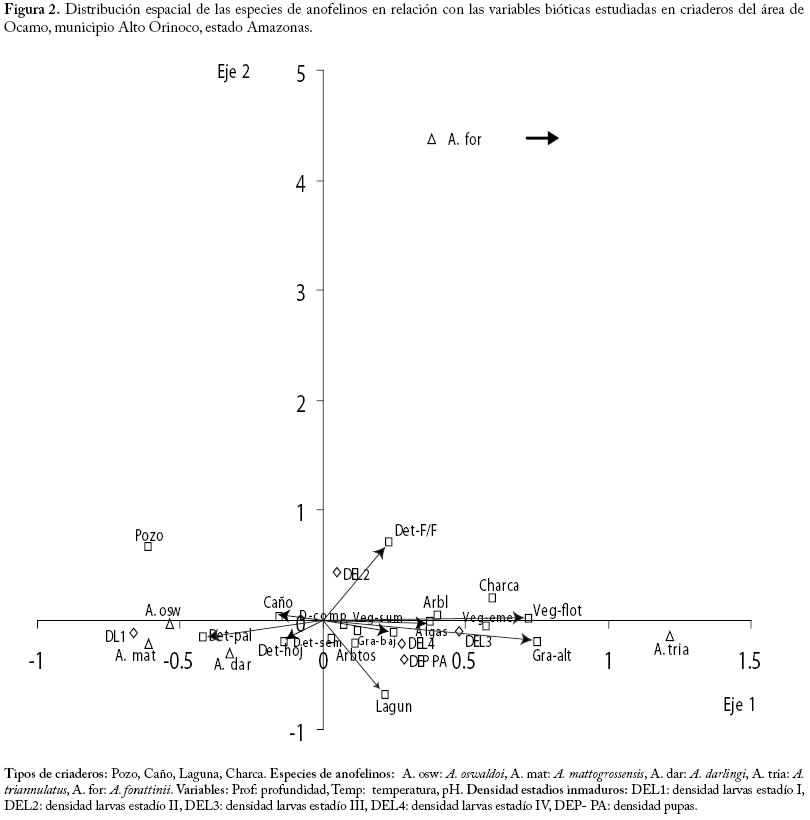
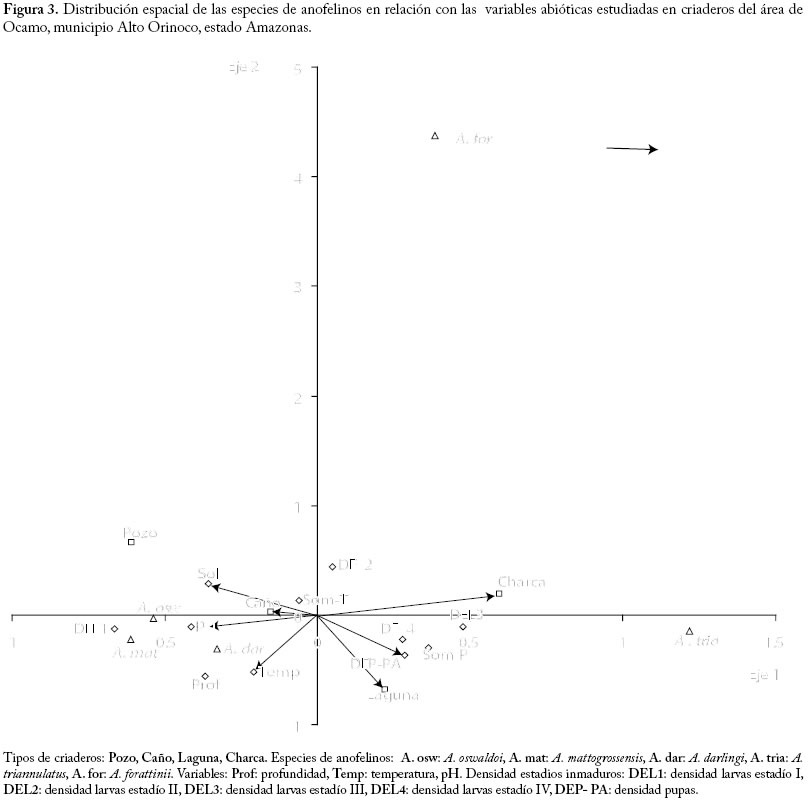
La especie más frecuente fue A. oswaldoi (61.3%), seguida de A. triannulatus (45.2%), A. darlingi (41.9%) y en proporciones menores se encontró A. mattogrossensis, A. punctimacula, A. forattinii, A. braziliensis y A. peryassui. De estas dos últimas se colectó un solo ejemplar (Cuadro 1). Del total de muestras tomadas (37), 83.8% resultaron positivas para larvas de anofelinos, estimándose la densidad en 0.89 larvas/cucharón (Cuadro 2) Pozo. Se localizó este tipo de criadero en lechos de caños y se forman en depresiones naturales durante la estación seca, quedando de esta manera estancados. Son temporales y cubiertos parcialmente de vegetación acuática (graminoides) y algas verdes filamentosas. Estos pozos estaban presentes en la selva con sombra parcial, generalmente de tamaño reducido (4 x 4 m), poco profundos, agua oscura, ligeramente turbia, con una temperatura promedio de 27,2 °C, pH de 6,0 y abundante detritus (Cuadro 3). La especie anofelina más abundante fue A. oswaldoi, mientras que solo se capturó un especímen de A. darlingi y A. forattinii (Cuadro 1). Este tipo de criadero presentó la mayor densidad larvaria (3.38 larvas/cucharón) en comparación con los demás tipos de criaderos (Cuadro 2). Caño. Se caracteriza por presentar corriente de agua, ancho y profundidad variable (Cuadro 3). El caudal de los caños dependerá de la época del año y de la intensidad de las lluvias. En relación al desplazamiento del agua, los criaderos evaluados tipo caño presentaron corriente leve, agua amarillenta con fondo fangoso, profundidad variable entre 60 centímetros y 3 metros. Estos caños generalmente servían de conexión entre lagunas y estaban profusamente sombreados. La temperatura promedio del agua fue de 26,1 °C y pH 6,3 (Cuadro 3). La densidad larvaria de este tipo de criadero fue la más baja, 0,13 larvas/cucharón (Cuadro 2). En este tipo de criadero se colectaron un total de 22 larvas, de las cuales solo fue posible identificar a nivel de especies cinco especimenes pertenecientes a las especies A. oswaldoi, A. forattinii y A. mattogrosensis (Cuadro 1). Charca. Este tipo de criadero tiene algo de similitud al pozo pero consideramos que por su reducido tamaño (1 x 1 metro) y por estar localizado en un lugar de paso de los indígenas Yanomami (camino en la selva), lo clasificamos como criadero tipo charca. Presenta alrededor de 10 cm de profundidad con abundante materia orgánica en descomposición, agua turbia por movimiento de lodo, sombra parcial, temperatura del agua de 32,2 °C y pH de 6,0 (Cuadro 3). La especie de anofelino más abundante en este tipo de criadero fue A. oswaldoi, seguido de A. triannulatus y A. darlingi en mínima proporción (Cuadro 1). La densidad estimada fue de 2,8 larvas/cucharón (Cuadro 2). Plantas acuáticas asociadas a criaderos de anofelinos colectadas. Para el estudio de la vegetación asociada a los criaderos sólo fue posible preservar e identificar aquellas plantas colectadas en los criaderos tipo laguna ubicados en los alrededores de los "shabonos" de Ocamo, Tumba, San Benito y Shashana. Las plantas provenientes de las lagunas cerca de Ocamo fueron identificadas como Salvinia auriculata (Salviniaceae), Elodea sp y Paspalum repens (Poaceae). Mientras que de la comunidad de Tumba se identificaron las siguientes especies: Elodea sp, Seleria seans (Cyperaceae) y Pistia stratiotes (Araceae); en San Benito se identificaron Elodea sp. y Heliconia psittacorum, mientras que en la laguna cerca de Shashana se colectó Panicun sp. (Poaceae), Salvinia auriculata (Salviniaceae) y Elodea sp. (Hydrocharitaceae). La única especie de planta colectada en todas las lagunas fue Elodea sp. Influencia de los factores bióticos y abiótico sobre la abundancia de especies. En las Figures. 2 y 3 se puede evidenciar que las variables bióticas detritus palitos (Det- pal) y detritus hojas (Det-hoj) y las variables abióticas (pH ácidos, profundidades de 1 a 3 m, y temperaturas entre de 26,7 °C a 28 °C), están asociadas positivamente con las especies más abundantes A. oswaldoi, A. darlingi y A. mattogrossensis. Con respecto a A. triannulatus, esta especie mostró preferencia por los hábitats tipo laguna con presencia de vegetación flotante y gramíneas altas. A. forattinii fue una especie muy poco abundante por lo tanto se comportó como una especie rara al no mostrar ningún tipo de asociación con las variables estudiadas. Por otra parte, las densidades de los estadios inmaduros de pupas (DEN-PA), larvas cuarto (DEL4) y tercer instar (DEL3), evidencian preferencia por las lagunas, ubicándose específicamente en sitios de sombra permanente y con abundantes algas. Los análisis de correspondencia canónica (ACC) realizados muestran que hay una fuerte estructuración en la relación entre las especies de anofelinos encontradas y las variables estudiadas. El ACC realizado para evidenciar la influencia de los factores bióticos (Figura 2) mostró que el segundo Eje explica 77% de la varianza, lo cual indica que existe una fuerte asociación entre las especies encontradas y las variables estudiadas, además estas variables sirven como indicadores para explicar la distribución espacial de las especies encontradas. Para re-evaluar este resultado se utilizó la prueba de permutación de Monte Carlo (Ter Braak I987a, b), lo cual permitió corroborar la existencia de una relación significativa (p=0,01) entre las especies de anofelinos colectadas y las variables ambientales estudiadas. Tipos de criaderos: Pozo, Caño, Laguna, Charca. Especies de anofelinos: A. osw: A. oswaldoi, A. mat: A. mattogrossensis, A. dar: A. darlingi, A. tria: A. triannulatus, A. for: A. forattinii. Variables: Prof: profundidad, Temp: temperatura, pH. Densidad estadios inmaduros: DEL1: densidad larvas estadío I, DEL2: densidad larvas estadío II, DEL3: densidad larvas estadío III, DEL4: densidad larvas estadío IV, DEP- PA: densidad pupas. Tipos de criaderos: Pozo, Caño, Laguna, Charca. Especies de anofelinos: A. osw: A. oswaldoi, A. mat: A. mattogrossensis, A. dar: A. darlingi, A. tria: A. triannulatus, A. for: A. forattinii. Variables: Prof: profundidad, Temp: temperatura, pH. Densidad estadios inmaduros: DEL1: densidad larvas estadío I, DEL2: densidad larvas estadío II, DEL3: densidad larvas estadío III, DEL4: densidad larvas estadío IV, DEP- PA: densidad pupas. Discusión Durante el presente estudio se identificaron un total de ocho especies de anofelinos, a saber A. oswaldoi, A. triannulatus, A. darlingi, A. mattogrossensis, A. punctimacula, A. forattinii, A. peryassui y A. braziliensis, lo cual contribuye a actualizar la lista de especies de anofelinos presentes en esta región (Moreno y Rubio-Palis 2003, Rubio-Palis 2005). Al comparar estos resultados con los de Rejmánková et al. (1999) hay algunas diferencias en cuanto a la diversidad de especies de anofelinos presentes en el área de estudio. En efecto, los dos estudios coinciden al reportar la presencia de A. triannulatus, A. darlingi, A. oswaldoi, A. punctimacula y A. peryassui. Es importante destacar que Rejmánková et al. (1999) reportan también la presencia de A. mediopunctatus, sin embargo estudios posteriores mostraron que esta especie había sido mal identificada y que en realidad se trata de A. forattinii (Moreno y Rubio-Palis 2003). El presente estudio reporta otras dos especies: A. mattogrossensis y A. braziliensis. Estudios similares realizados en el estado Bolívar reportan un número mayor de especies de anofelinos (12), además de Chagasia bathana, género perteneciente a la tribu Anofelini (Moreno et al. 2000). Estos resultados parecen confirmar los hallazgos de Tadei et al. (1998) en la Amazonía brasilera, donde la diversidad de especies de anofelinos es mayor en ambientes alterados por el hombre debido al incremento de nutrientes en los criaderos causado por las actividades agrícolas. En el área de Ocamo se identificaron cuatro tipos de criaderos: lagunas, pozos, caños y charca. Las lagunas, al igual que en otros estudios realizados en Venezuela (Moreno et al. 2000, Rejmánková et al. 1999), resultaron los hábitats más favorables para el desarrollo de diversas especies de anofelinos, posiblemente debido a que constituyen un hábitat permanente y la presencia de abundante vegetación flotante y emergente puede producir un abundante perifiton que suministra alimento para las larvas (Merritt et al. 1992), así como también la abundancia de vegetación provee de refugios que protegen a las larvas de la depredación (Hall 1972). La especie más abundante y frecuente durante este estudio fue A. oswaldoi, encontrándose en los cuatro tipos de criaderos y durante los cuatro meses que duró el estudio de campo. Esta especie fue colectada en sitios de poca profundidad y sombra parcial, prefiriendo los criaderos tipo pozo y caño (Figs. 2 y 3). Estos resultados contrastan con lo reportado por Moreno et al. (2000) en el estado Bolívar y Grillet et al. (1998) en el estado Sucre, quienes señalan para esta especie a las lagunas como los criaderos preferibles; posiblemente esto sea debido a que en el presente estudio se realizó un análisis (Correspondencia Canónica) mucho más preciso (CANOCO). La segunda especie colectada con mayor frecuencia fue A. triannulatus, la cual mostró preferencia por aguas profundas y criaderos tipo laguna con vegetación flotante y gramíneas altas (Figs. 2 y 3), coincidiendo con los resultados reportados por Rejmánková et al. (1999). Sin embargo, es importante señalar que en los estudios realizados por estos autores durante el pico de lluvias en Julio de 1997, la especie más abundante y frecuente en las lagunas fue A. triannulatus mientras que sólo se colectó un espécimen de A. oswaldoi. En estudios realizados en diversos tipos de criaderos en áreas maláricas del estado Bolívar, reportan que la especie más abundante y con más amplia distribución es A. triannulatus (Berti et al. 1998, Moreno et al. 2000), sin embargo esta especie no parece estar involucrada en la transmisión de malaria (Moreno et al. 2000). Esta podría ser la situacuón en el Alto Orinoco, donde A. triannulatus sólo ha sido colectada sobre cebos humanos ubicados próximos a los criaderos entre las 1800 y 2000 horas (Rubio-Palis et al. datos no publicados). Anopheles darlingi, es el vector regional de malaria en el Alto Orinoco y representa el 99% de los anofelinos capturados con cebos humanos y trampas de luz dentro de las viviendas (Rubio-Palis 1995, Rubio-Palis et al. 2000). La baja abundancia de A. darlingi encontrada durante este estudio parece coincidir con reportes previos donde se señala una reducción en la abundancia de los adultos hacia el final del período de lluvias e inicio de la sequía (Magris et al. 1999, Rubio-Palis et al. 2000). A. darlingi fue colectada en mayor proporción en el criadero tipo laguna, pero también se le colectó en los criaderos tipos pozo y charca, ubicados en lugares con poca profundidad, sombra parcial y abundante detritus (palitos, hojas, semillas) asociada con A. oswaldoi (Figura 2). Lo antes expuesto concuerda con la descripción de Fleming (1986), en donde manifiesta que las larvas de A. darlingi ocupan una gran diversidad de hábitats, pero siempre en agua dulce, no contaminada y con sombra parcial, es decir, protegidos de los rayos directos del sol al menos durante parte del día. Mientras que en los ríos de Belice, Rejmánková et al. (2000) reportan a A. darlingi asociada a ensamblajes de detritus. Anopheles mattogrossensis, también fue colectada durante los cuatro meses de evaluación, pero en proporciones menores. Esta especie se colectó principalmente en criaderos tipo laguna y una sola vez en un caño en un lugar sombreado y poco profundo (Cuadro 1). Anopheles forattinii se colectó en un lugar sumamente sombreado por árboles emergentes de gran altura en donde abundaban macrofitas (Heliconia spp.) y con abundante detritus (palitos, resto de flores y frutas flotando), en camino anegado en dirección a una laguna; también esta especie se colectó en criaderos tipo caño, con poca profundidad, sombreado, agua cristalina con fondo limoso y escaso detritus (palitos y flores). Este reporte constituye un aporte significativo al conocimiento de esta especie, ya que ha sido descrita recientemente (Wilkerson y Sallum 1999) y poco se conoce sobre su distribución geográfica y bionomía. En general, nuestros resultados contrastan con los reportes de Rejmánková et al. (1992, 1993, 1998, 2000) en México y Belice para A. albimanus, A. pseudopunctipennis, A. vestitipennis y A. punctimacula, donde la presencia de larvas de anofelinos es controlada por la vegetación acuática dominante y no por las características físicas de los criaderos como hidrografía y profundidad. El presente trabajo puede servir de base para nuevos estudios sobre bionomía y ecología de los estadios inmaduros de anofelinos, ya que los resultados obtenidos sugieren que a fin de caracterizar adecuadamente la riqueza de especies y abundancia de anofelinos en criaderos del Alto Orinoco deberían hacerse estudios longitudinales a fin de realizar colectas mensualmente a lo largo de al menos 15 meses. Este tipo de estudios son fundamentales para el diseño, planificación e implementación de medidas de control efectivas. Agradecimientos A Jorge E. Moreno y Alfredo Gutiérrez por la ayuda brindada en el análisis de los datos, a Teresa Méndez por la producción del mapa del área de estudio. Al árbitro por sus críticas y sugerencias. Este trabajo fue posible gracias al financiamiento del Proyecto Control de Enfermedades Endémicas (PCEE-021-002). Referencias
© 2005- Sociedad Venezolana de Entomología The following images related to this document are available:Photo images[em05006f3.jpg] [em05006f2.jpg] |
| |||||||||
{kind=link}
{kind=link}
{kind=link}