 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Entomotropica antes/formerly Boletín de Entomología Venezolana, Vol. 20, No. 1, April, 2005, pp. 39-47 Distribución de Brevipalpus phoenicis (Geijskes) (Acari: Tenuipalpidae) en la planta del guayabo (Psidium guajava L.) en La Coruba, municipio Mara, estado Zulia, Venezuela Distribution of Brevipalpus phoenicis (Geijskes) (Acari: Tenuipalpidae) on guava plant (Psidium guajava L.) in La Coruba, Mara county, Zulia state, Venezuela Magally Quirós, Yadira Petit, Nedy Poleo y Ángel Gómez Universidad del Zulia, Ciudad Universitaria, Facultad de Agronomía, Programa Museo de Artrópodos, MALUZ. CC-0133-02, Maracaibo, Zulia, Venezuela. mquiros@cantv.net, yadirapetit@yahoo.com, nedypoleo@hotmail.com, gomezyasociados5@cantv.net Code Number: em05007Resumen QUIROZ M, PETIT Y, POLEO N, GÓMEZ A. 2005. Distribución de Brevipalpus phoenicis (Geijskes) (Acari: Tenuipalpidae) en la planta del guayabo (Psidium guajava L.) en La Coruba, municipio Mara, estado Zulia, Venezuela. Entomotropica 20(1): 39-47. Para determinar la distribución de B. phoenicis en plantas de guayabo y especificar la unidad de muestreo que refleje de manera confiable la infestación de esta plaga, se realizó un estudio durante 9 meses, en plantas de 7 años en La Coruba (lat 121º46’75’’N; long 18º15’91’’E). La copa de la planta se dividió en cuatro cuadrantes (norte, sur, este, oeste) y verticalmente en estratos basal (EB) y apical (EA). Se contaron en el laboratorio las formas móviles del ácaro en 4 hojas en desarrollo (HD), 4 hojas plenamente desarrolladas (HPD) y 4 frutos verdes (FV) muestreados de cada ubicación, para un total de 384 órganos/mes. Las medias de B. phoenicis fluctuaron significativamente (P<0,01) según las fechas, los valores más altos se registraron en junio, agosto y septiembre de 2002. Las medias de B. phoenicis más altas (P<0,01) se distribuyeron en los FV (4,53) seguidos de las HPD (1,49) y HD (0,80). No hubo diferencias entre medias del B. phoenicis en los cuadrantes, pero la media máxima se registró para el norte (2,64). Tampoco hubo diferencias entre las medias de B. phoenicis distribuidos en los EB y EA (P>0,05), pero hubo diferencias (P<0,01) en las medias de las interacciones pares entre fecha y cada uno de los factores principales, así como entre cuadrantes × estratos. B. phoenicis se distribuye en toda la copa y el muestreo de los frutos en cualquier cuadrante y estrato estimaría de manera confiable la abundancia del ácaro durante el desarrollo de los frutos. Esta información sirve para desarrollar un plan de muestreo de presencia/ausencia para estimar la proporción de FV infestados, ayudando a la toma de decisiones del manejo en la zona de estudio. Palabras clave adicionales: Ácaro fitófago, ácaro plano, dispersión, unidad de muestreo. Abstract QUIROZ M, PETIT Y, POLEO N, GÓMEZ A. 2005. Distribution of Brevipalpus phoenicis (Geijskes) (Acari: Tenuipalpidae) on guava plant (Psidium guajava L.) in La Coruba, Mara county, Zulia state, Venezuela. Entomotropica 20(1): 39-47. A study was conducted during 9 months period, to get the within-plant distribution of B.phoenicis and to determine the sampling unit that would estimate infestation on 7 year-old guava plants at La Coruba (Lat. 121º46’75’’N Long. 18º15’91’’E). The canopy of the plant was divided into four quadrants: north, south, east, and west, and vertically in basal (BS) and apical strata (AS). Motile forms of the mite were counted from 4 young leaves (YL), 4 physiologically mature leaves (PML) and 4 green or unripe fruits (GF) picked up from each quadrant of the canopy. A total of 384 leaves and fruits were selected monthly. The means fluctuated significantly (P<0.01) when considering sampling dates, highest values registered for June, August, and September of 2002. The highest means of B. phoenicis (P<0.01) were distributed on the GF (4.53) followed by PML (1.49) and YL (0.80). There were no significant differences among the means of B. phoenicis on the quadrants, but the maximum value was observed on the north (2.64). There were no significant differences among the means on the vertical strata (P>0.05), but there were differences (P<0.01) among the means, considering the interactions between the date x each individual factor, as well as between the quadrant × strata interaction. B. phoenicis is widely distributed on the canopy of the 7 year-old plants. The GF sampled from any strata and quadrant would give a reliable estimation of the mite density during the fruit growth. This information is important when developing a presence/absence sampling program to estimate the mite infestation faster and easily, thus helping on the decision making process to manage this pest. Additional key words: Dispersion, flat mite, plant-feeding mite, sampling unit. Introducción A nivel mundial el guayabo es uno de los tantos hospederos del ácaro plano, Brevipalpus phoenicis (Geijskes) (Jeppson et al. 1975), causa daños leves a severos de manera localizada en el ápice, base y pedúnculo de los frutos y ocasionalmente sus poblaciones son tan altas que dañan todo el pericarpio del fruto (Guerere y Quirós 2000; Quirós et al. 2002); también habita y se alimenta de la savia del follaje, ramas, botones y flores de las plantas (Quirós y Viloria 1991; Camacho et al. 2002). En el Zulia, todas las plantaciones visitadas de este frutal se encuentran infestadas por esta especie (información tomada de la Base de Datos del Programa financiado por CONDES, LUZ #CC-0133-02), haciéndose más notables los daños en las épocas de sequías prolongadas, por lo que los productores utilizan acaricidas comerciales para suprimir las poblaciones de manera curativa y sin conocimiento de los niveles de infestación del ácaro, es decir cuando los daños están avanzados en los frutos, traduciéndose en altos costos para el productor debido a que pudieran estar tratando tardíamente o en otros casos, pudieran estar eliminando los insectos o ácaros beneficiosos que depredan sobre esta u otra plaga del cultivo. Hoy en día, es necesario desarrollar la información biológica, ecológica y etológica de las especies fitófagas claves que se presentan en las diferentes regiones, así como bajo las diversas condiciones de manejos agronómicos para poder idear e implementar un manejo de plagas integral del cultivo. Uno de los tantos aspectos consiste en el desarrollo de un plan de muestreo que estime las poblaciones de la misma en un límite de tiempo razonable (Mollet et al. 1984), sin embargo para poder lograr esto se debe tener información de cómo se distribuye la misma en la planta, a fin de poder seleccionar las muestras que aporten una estimación confiable de la densidad o de la presencia de la plaga (Opit et al. 2003). Las características de las plantas, entre otras la calidad nutritiva es un factor clave determinante en la fecundidad y en las estrategias de reproducción (Ej.: tamaño y calidad de los huevos, la distribución de recursos para los huevos, la escogencia del sitio de oviposición) de las especies fitófagas, lo cual a su vez, influye en la abundancia y distribución en la planta de las mismas. Las respuestas de las especies fitófagas a los cambios de calidad del hospedero varían dentro y entre los sitios de alimentación en la planta. El hospedero también puede afectar interacciones de alto nivel trófico (del individuo y de las poblaciones) tales como el desenvolvimiento de los depredadores y parasitoides cuando responden a estímulos químicos volátiles que producen las plantas cuando son atacadas por los herbívoros o cuando estos oviponen sobre el tejido vegetal (Awmack 2002; Hilker et al. 2002), inclusive se han reportado señales de stress emitidas por plantas atacadas por artrópodos a plantas sanas (Pickett 2003) y también se han reportado modificaciones fenotípicas como mecanismos de defensa que inducen a la resistencia o susceptibilidad de la planta a los insectos herbívoros (Van Zandt y Agrawal 2004). La plantación escogida para el presente estudio tuvo características muy particulares de conformación de la copa, la cual puede definirse como bastante uniforme, de porte alto y buena frondosidad, a diferencia de las plantaciones de guayabo de la zona tradicional del municipio Mara, las cuales tienen copa de arquitectura irregular, de porte bajo y poco frondosas. Esto podría tener cierta influencia, entre otros factores, en la forma de distribución en la planta de las poblaciones de B. phoenicis. En todo programa de manejo de plagas se hace necesario conocer donde se encuentra la plaga debido a la preferencia de esta por un determinado órgano o por la susceptibilidad de la planta a ser afectada en mayor proporción en algunas de sus estructuras en algún momento determinado. El presente estudio tuvo como objetivo determinar la distribución del ácaro B. phoenicis en la copa de la planta y especificar la unidad de muestreo u órgano de la planta que refleje de manera confiable la infestación de esta plaga. Materiales y Métodos El estudio se llevó a cabo en la granja La Coruba ubicada en el municipio Mara, estado Zulia, Venezuela (Lat. 121º46’75’’N Long. 18º15’91’’E), desde junio 2002 a marzo 2003. La precipitación promedio anual de la zona es 800 mm (Estación Meteorológica Carrasquero, Municipio Mara, Zulia, Venezuela), distribuidos en dos picos desiguales en los meses de mayo y octubre, siendo este último el más abundante. La temperatura promedio anual es 28ºC y la humedad relativa promedio es 75%. Estas condiciones determinan a su vez los picos de producción que se expresan en dos períodos bien definidos durante el año, donde la precipitación bimodal es factor clave en las dos emergencias florales de considerable magnitud, que se traducen en dos épocas de cosecha durante el año, uno entre los meses de diciembre, enero y febrero y el otro de menor magnitud en junio, julio y agosto (Tong et al. 1991; Quijada et al. 1999). Las plantas de guayabo utilizadas fueron del tipo Criolla Roja, de 7 años de edad. Las copas de esas plantas se describen como: redondeadas, ramificadas internamente, de tamaño y forma homogénea; sembradas a una distancia de siembra de 7 x 7 e irrigadas por un sistema de platones, con agua proveniente del río Socuy. Los materiales y equipos utilizados fueron: bolsas de papel, bolsas plásticas, cavas conservadoras, portaobjetos, medio de montaje Hoyer, estereoscopios LEICA® . Los muestreos se hicieron en 4 plantas escogidas al azar para un total de 36 plantas y realizados en el siguiente orden cronológico: 19-06-2002, 01-07-2002, 02-08-2002, 11-09-2002, 09-10-2002, 04-11-2002, 22-01-2003, 04-02-2003 y 24-03-2003. La copa de la planta se dividió en los cuadrantes norte, sur, este, oeste y cada cuadrante en dos estratos verticales: estrato apical (EA), por encima de 1,50 m de altura y estrato basal (EB) por debajo de 1,50 m. Como no se tenían antecedentes para este cultivo y plaga, sobre la preferencia del ácaro por un órgano en particular, las unidades de muestreo fueron seleccionadas al azar, considerando 3 tipos de órganos con características diferentes, como se indica a continuación: 4 hojas en desarrollo (HD) ubicadas en el segundo entrenudo de las ramas de la planta (en dirección ápice-base), de textura suave, flexible, pubescente y coloración verde claro; 4 hojas plenamente desarrolladas (HPD) del sexto entrenudo de las ramas, de textura dura, glabras, quebradizas y coloración verde oscuro; y 4 frutos verdes (FV). Esos tres tipos de órganos tienen características fisiológicas, morfológicas y fenológicas diferentes (C. Fernández B., Fisiólogo, Dpto. Botánica, Facultad de Agronomía, LUZ, Comunicación Personal), lo cual a su vez influiría sobre la distribución de las poblaciones de este ácaro fitófago. Estas hojas y frutos fueron tomados de cada estrato vertical dentro de cada cuadrante de la copa de la planta. El muestreo en el campo fue realizado por cuatro personas de estaturas entre 1,54 m y 1,60 m. Un total de 3 456 órganos fueron muestreados, distribuidos en 1 152 muestras para cada tipo de órgano, o sea 384 órganos/fecha. Las muestras se llevaron al laboratorio en bolsas debidamente identificadas, trasladadas y mantenidas en las cavas por uno o dos días, correspondientes a la observación y conteo de las formas móviles del ácaro bajo los estereoscopios, los cuales fueron realizados por tres personas. Simultáneamente se montaron en portaobjetos en medio de Hoyer algunos especimenes para cada fecha de muestreo, los cuales se depositaron en la colección del Museo de Artrópodos de LUZ (MALUZ). El diseño utilizado fue un diseño no experimental, exposfacto longitudinal de campo, donde se estudiaron las relaciones entre variables, considerándose en este caso como variables antecedentes: fecha, tipo de órgano, cuadrante y estrato en el cuadrante y como variable respuesta: el número de ácaros. Las unidades de muestreo fueron HD, HPD y FV. Se uso el programa Statistical Analysis System (SAS, 2000), efectuándose el análisis de varianza (ANOVA) mediante el procedimiento PROC GLM, tomando en cuenta el efecto de las variables antecedentes sobre el número de ácaros/órgano, la prueba de comparaciones múltiples de medias de Tukey en los efectos principales significativos y el procedimiento LSMEANS para los efectos múltiples de las interacciones significativas. Se utilizaron tablas cruzadas de distribución de frecuencia entre: fecha y tipo de órgano, entre fecha y cuadrante, entre fecha y estrato y entre cuadrante y estrato para estimar el porcentaje de ácaros para cada celda; el estadístico Chi-cuadrado de Pearson (χ2) fue aplicado para determinar la asociación entre las variables. Como el número de ácaros no presentó una distribución cercana a la normal, se utilizó la transformación logarítmica: Log (número de ácaros + 10). Se presentan figuras que ilustran la fluctuación poblacional de B. phoenicis, su distribución en los diferentes tipos de órganos, cuadrantes, así como el comportamiento de las medias según fechas y tipos de órganos y la asociación entre medias según los estratos y los cuadrantes de las plantas. Resultados y Discusión Según el ANOVA (Cuadro 1) la distribución de B. phoenicis en la planta del guayabo estuvo afectada en forma altamente significativa (P<0.01) por los factores principales estudiados: fechas de muestreo y tipo de órgano, más no por los cuadrantes ni por los estratos verticales de la copa de la planta. Mientras que las interacciones estadísticas entre pares de factores principales fueron altamente significativas (P<0,01) para: fechas x tipos de órgano; fechas x cuadrantes; fechas x estratos; cuadrantes x estratos de la copa de la planta, indicando que la variabilidad del número de ácaros fue explicada por la fecha de muestreo, tipo de órgano, estratos y cuadrantes en el período considerado en el estudio. Cuadro 1. Análisis de varianza para el número de B. phoenicis según su distribución en la planta del guayabo en la Coruba, municipio Mara, estado Zulia, Venezuela.
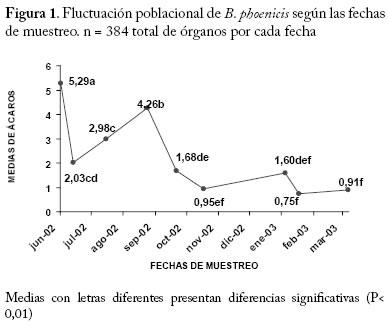
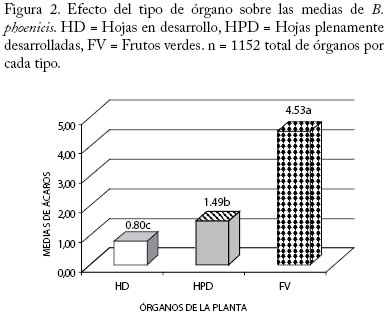
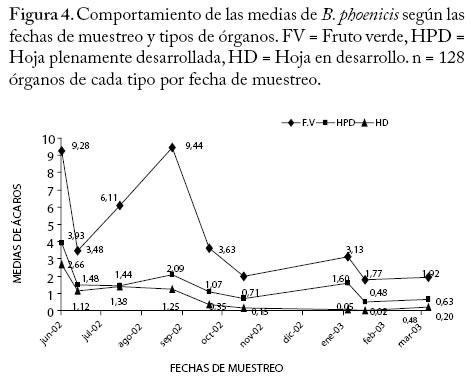
** Diferencias altamente significativas (P<0.01) CV = 9,97 % * Diferencias significativas (P<0.05) Efecto Fecha sobre el número de B. phoenicis. En la Figura 1 se muestra el comportamiento de las medias de B. phoenicis en las diferentes fechas de muestreo, indicando que la densidad de B. phoenicis cambió con las fechas. Se observó una fluctuación en la población del ácaro en el período de realización de este estudio, cuyas medias más altas (P<0.01) se presentaron en los meses de junio (5,29), agosto (2,98) y septiembre (4,26) del 2002, mientras que las medias más bajas se encontraron en julio-Agosto y desde octubre 2002 hasta marzo del 2003. Estos efectos diferenciales en la distribución y abundancia del ácaro con respecto al tiempo pudo deberse a un conjunto de factores entre las cuales por ejemplo se citan a las condiciones climáticas, estudios previos en otros cultivos señalan que esas condiciones influyen en la distribución dentro de la planta de los artrópodos herbívoros y de sus depredadores (González et al. 2002), mientras que otros enfatizan que la precipitación es un factor extremadamente limitante en la dinámica poblacional de los ácaros fitófagos, además de que seria simplista no considerar que muchos de los factores físicos y ambientales interactúan influenciando sobre la probabilidad de aparición de los ácaros fitófagos como plagas (Gray et al. 1988). En La Coruba la aplicación de productos químicos insecticidas y acaricidas para controlar Capulinia sp., Liothrips sp. y B. phoenicis fue bastante frecuente, lo cual pudo tener influencia en ese comportamiento de las poblaciones del ácaro plano. Se ha determinado que ocurren cambios en la distribución de los ácaros depredadores en plantas de manzana como consecuencia de la aplicación de productos químicos (Fitzgerald y Solomon 2002). Por otro lado, la fluctuación poblacional de B. phoenicis pudo depender también de los cambios en la morfología y fisiología de la planta como consecuencia del manejo agronómico empleado y de los artrópodos plagas presentes en el cultivo. Se ha reportado que la calidad de la planta, en términos del carbono, nitrógeno y metabolitos de defensa, es factor determinante de la fecundidad real o potencial de los artrópodos herbívoros y depredadores de manera individual y a nivel de poblaciones (Awmack y Leather 2002), lo cual también pudo haber afectado a favor o en contra de los depredadores de B. phoenicis. Estudios realizados en otros cultivos indican que la edad de la planta y los cambios fenológicos que pudieran tener relación con la pérdida de follaje, bien sea por senescencia o por daños de artrópodos también pudiera afectar la distribución dentro de la planta de las plagas debido al cambio en el microclima de ciertas partes de las planta que inducen a la movilización de un sitio a otro de las mismas a lo largo del tiempo (Maisin et al. 1997). Efecto del tipo de órgano sobre B. phoenicis. Según la prueba de comparaciones múltiples de Tukey la media más alta de ácaros se encontró en los FV (4,53) en comparación con las medias de las HPD (1,49) y de las HD (0,80) (P<0,01) (Figura 2). Según estudios previos B. phoenicis invade al fruto desde su formación, afectando primero al pedúnculo y base y posteriormente al ápice y sépalos, permaneciendo las diferentes generaciones durante todo el crecimiento del fruto (Quirós et al. 2002). Esta especie por su cuerpo característicamente plano se encuentra con frecuencia en las depresiones y hendiduras del tejido apical y basal del fruto, así como en las caras internas y externas de los sépalos, lo cual es probablemente una estrategia de defensa del ácaro contra sus enemigos naturales o una estrategia de selección del alimento (nutrientes) en esos sitios de preferencia como respuesta a algún tipo de estímulo proveniente de la planta. Brevipalpus phoenicis, también se localiza en el envés de la hoja, especialmente a lo largo de las nervaduras de las HPD y de las HD, en las cuales hay menos variabilidad de estructuras que puedan servir de refugio a las poblaciones. Numerosos estudios se han realizado en otras especies que tratan este tipo de interacción planta y ácaro fitófago que explican la selectividad del hospedero, así como de partes u órganos de este. Esas interacciones a favor de un hospedero en especial, es uno de los factores que determina que ciertos artrópodos prefieran más a una especie de planta que a otra (Gould 1978; Fox y Morrow 1981; Futuyma y Moreno 1988; Jaenike 1990; Agrawal 2000). Efecto del cuadrante de la copa de la planta sobre B. phoenicis. En la Figura 3, se muestra que no hubo efecto significativo (P>0.05) del cuadrante (norte, sur, este y oeste) sobre el número de B. phoenicis, sin embargo se observa que estas oscilaron entre 2,64 y 2,03, siendo el valor máximo para el cuadrante norte de la copa de la planta. El guayabo es un cultivo de ciclo largo con una vida productiva que no debería exceder los 8 años de edad (Morales y Rodríguez 2002), la plantación de La Coruba para el momento del estudio tenía 7 años, lo cual pudiera estar relacionado con la dispersión y permanencia de esta plaga en todos los lados de la planta. Esta especie se moviliza desde las hojas viejas y ramas colonizando los tejidos nuevos vegetativos, brotes florales o futuros frutos, dependiendo de su evolución fenológica (Quirós et al. 2002). Debido a que la planta siempre tiene frutos y hojas plenamente desarrolladas esta especie se ve favorecida en su permanencia en la planta. Se ha observado que las plantas en algunos guayabales del municipio Mara tienen copas redondeadas o por lo contrario, dirigidas hacia una dirección determinada debido a los efectos del viento sobre algunos de los cuadrantes (Villavicencio et al. 1995). En La Coruba, las condiciones edafoclimáticas y el manejo del riego por platón de agua proveniente del Río Socuy quizás ha tenido su influencia sobre la forma de las copas de las plantas las cuales son bastantes uniformes, redondeadas y frondosas a pesar de la edad, lo cual probablemente también tuvo relación con el comportamiento de distribución del ácaro muy similar en todos los cuadrantes. Interacciones El ANOVA muestra que las interacciones entre fecha y cada uno de los factores principales tipo de órgano, cuadrante y estrato afectó significativamente (P<0,01) a las medias poblacionales de B. phoenicis (Cuadro 1). En los Cuadros 2, 3, 4 se muestra la distribución porcentual del ácaro para cada interacción y según la prueba de Chi-cuadrado de Pearson los valores de χ2 para órganos (FV, HPD, HD), cuadrantes (norte, sur, este y oeste) y estratos (EB y EA) fueron 357,29, 509,75 y 262,44, respectivamente (P<0,0001). A continuación se analizan cada una de las interacciones antes señaladas. Interacción Fecha × Órgano El número de ácaros en los 3 tipos de órganos fluctuó con tendencias similares en cuanto a los ascensos y descensos a lo largo del período estudiado (Figura 4). Aparentemente los mismos factores bióticos y abióticos, así como las subsecuentes interacciones entre ellos determinaron este comportamiento en los frutos y hojas. Entre las medias del ácaro en los FV y HD se observaron diferencias altamente significativas (P<0.01) para la mayoría de las fechas. Mientras que entre los FV y HPD hubo diferencias significativas solamente en el período junio a octubre del 2002 (Figura 4). Las medias en las HPD estuvieron por encima de las medias de ácaros en las HD, sin embargo no hubo diferencias significativas entre ellas (P>0,05), lo que indica que la distribución del ácaro fue muy parecida en ambos tipos de hojas en el período del estudio. Considerando los tres tipos de órganos en conjunto, las medias oscilaron entre 0,75 (febrero 2003) y 5,29 (junio 2002) (Figura 1). Para cada tipo de órgano por separado, los ácaros se distribuyeron en mayor número en los FV, cuyas medias oscilaron entre 1,92 y 9,28 con respecto a las fechas del estudio. En las HPD las medias del ácaro oscilaron entre 0,48 y 3,93. Es decir, la distribución del ácaro en los órganos fue de un 66,43 % en los FV; 21,87 % en las HPD y 11,70 % en las HD (Cuadro 2). Los meses más críticos fueron junio (inicio del primer pico de cosecha) y septiembre 2002 (período de recuperación y reposo de las plantas para dar inicio al segundo pico de floración), ya que los frutos presentaron mayor abundancia del ácaro con respecto a las otras fechas (15,13 % y 15,38 % respectivamente), igual tendencia se observó en las HPD (Cuadro 2). Los cambios fenológicos ocurridos en las plantas para esos dos ciclos de producción que comprendió el estudio y los cambios climáticos, principalmente los períodos prolongados sin precipitaciones probablemente influyeron en ese comportamiento. Igualmente en estudios previos sobre las plantaciones existentes en el municipio Mara, han determinado que existe una gran variabilidad genética y en consecuencia una marcada variación en las características químicas y físicas de los frutos, las que también dependen del manejo agronómico de la plantación, de la época de cosecha y del estado de madurez de los frutos (Tong et al. 1991; Marín et al. 1993), lo cual podría haber tenido una incidencia sobre las poblaciones del ácaro. Cuadro 2. Distribución porcentual de B. phoenicis por fecha de muestreo en los diferentes tipos de órganos. FV = Fruto verde HPD = Hoja plenamente desarrollada HD = Hoja en desarrollo.
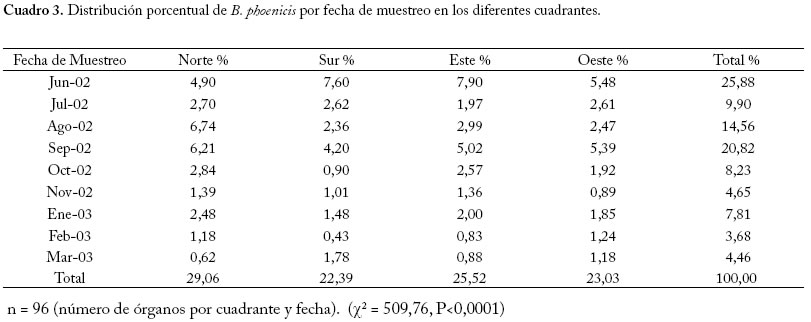
n = 128 (número de órganos por fecha). (χ2 = 357,30, P<0,0001) Interacción Fecha × Cuadrante El número de ácaros considerando los cuadrantes varió con las fechas, oscilando entre 6,46 en el cuadrante este (junio 2002) y 0,35 en el cuadrante sur (febrero 2003), no se observó un patrón definido en cuanto a la preferencia de las poblaciones por un cuadrante en particular, sin embargo en el cuadrante norte se presentaron con mayor frecuencia (en 6 fechas continuas) medias máximas del ácaro desde el mes de julio 2002 hasta enero 2003 (Figura 5). En el Cuadro 3 se muestran las proporciones del ácaro por cuadrantes con respecto a las fechas de muestreo. El primer ciclo de producción de frutos del año 2002 en comparación con el segundo ciclo, tuvo mayores proporciones del ácaro, afectando a la planta en todos los cuadrantes desde junio a septiembre. Para el desarrollo de un protocolo de muestreo se recomienda considerar las diferencias en los patrones poblacionales determinadas según las épocas de producción (Reitz 2002). En el municipio Mara algunas plantaciones han mostrado en sus copas el efecto del viento, haciendo que las ramas expuestas a este factor climático se desarrollen menos por las altas tasas de transpiración que este provoca (Villavicencio et al. 1995), este efecto se presenta más a principios del año en la zona de estudio. Las copas de las plantas seleccionadas para el estudio no estuvieron deformadas por el viento y sus copas eran redondeadas y más uniformes lo cual quizás favoreció la distribución del ácaro en todos los cuadrantes sin diferencias significativas entre ellos. Cuadro 3. Distribución porcentual de B. phoenicis por fecha de muestreo en los diferentes cuadrantes.
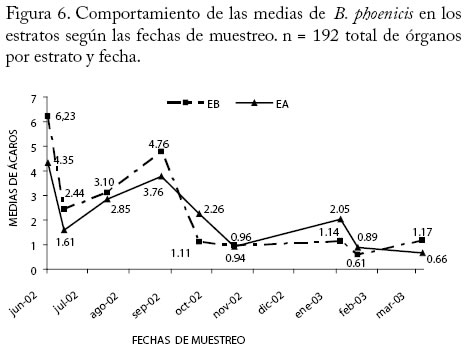
n = 96 (número de órganos por cuadrante y fecha). (χ2 = 509,76, P<0,0001) Interacción Fechas×Estratos verticales En la Figura 6 se muestra el comportamiento de las medias de B. phoenicis en los estratos verticales de la planta (EB y EA) según las fechas de muestreo. Entre los meses de junio a septiembre de 2002 y marzo del 2003, las medias fueron más altas en el EB mientras que de octubre 2002 a febrero del 2003 ocurrió así en el EA. Las proporciones del ácaro en los estratos basal y apical variaron según las fechas de muestreo, en 6 de las 9 fechas se encontraron valores máximos del ácaro en el estrato basal de la copa (Cuadro 4). Estudios sobre la producción de guayaba en la Planicie de Maracaibo indican que la mayor producción de frutos de este frutal está localizada en la parte media y basal de la copa (M. Marín, Fisióloga, Dpto. Botánica, Facultad de Agronomía, LUZ, Comunicación Personal) lo cual explica la abundancia del ácaro en ese estrato de la copa con mayor frecuencia en dependencia con la fecha. Considerando la forma y tamaño de B. phoenicis, así como su vida estratégica en cuanto a su localización en los numerosos habitas en la planta esta especie permanece en ella, dispersándose las poblaciones a medida que se forman nuevas ramas y en consecuencia frutos con el avance en edad de la planta. Estudio relacionado con la arquitectura de la planta de algodón según su estado de desarrollo determinó que una mayor densidad de hojas pudo alterar el microclima en la copa de la planta y por lo tanto afectó indirectamente los estratos que los insectos ocupaban (Atakan et al. 1996). Cuadro 4. Distribución porcentual de B. phoenicis por fecha de muestreo en los estratos basal (EB) y apical (EA).
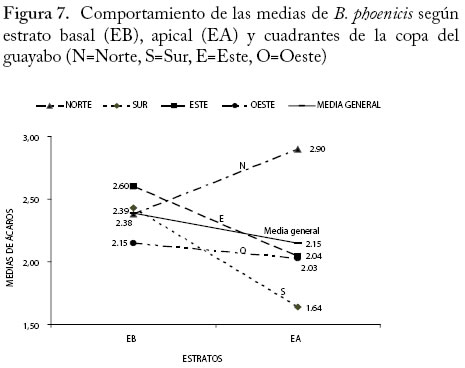
n = 192 (número de órganos por estrato y fecha). Interacción Cuadrantes × Estratos verticales En la Figura 7 se observa que las medias de B. phoenicis en los cuadrantes este, sur y oeste fueron más altas en el EB que en el EA. Mientras que en el cuadrante norte se presentó la media más alta en el EA, además de que la media del ácaro en el EB del mismo cuadrante estuvo muy cercana a los otros cuadrantes y a la media general. La prueba de Chi-cuadrado de Pearson (χ2) encontró que la interacción entre los estratos verticales (EB y EA) y los cuadrantes de la planta son dependientes, con una significancia del 5% con respecto a las medias de B. phoenicis, es decir el comportamiento de las medias del ácaro en los estratos no fue el mismo en todos los cuadrantes. Trabajos similares sobre distribución dentro de la planta pero con otra especie de ácaro señalan que la abundancia de Trisetacus juniperinus en los brotes apicales y subapicales del estrato superior del Cupressus sempervirens depende de las características fisiológicas y del estatus nutricional del tejido del cual se alimenta el ácaro (Castganoli y Simoni 2000). Conclusiones Se concluye que para las condiciones del período del estudio, en plantas de 7 años B. phoenicis se distribuye en todos los cuadrantes y estratos, por lo tanto el muestreo de los frutos verdes en cualquier parte de la copa estimaría mejor y de manera significativa, si se compara con las hojas, la abundancia relativa del ácaro durante el desarrollo de los frutos. Esta información servirá de base para desarrollar en un futuro próximo un plan de muestreo presencia-ausencia que permita estimar la proporción de frutos verdes infestados de manera rápida, ayudando a la toma de decisiones del manejo de la misma en la zona de estudio. Sería recomendable realizar estudios de comparación entre fincas de la misma y otras zonas de producción, así como de plantaciones más jóvenes. Se concluye también que el ácaro plano, B. phoenicis, varía en su distribución y abundancia en la planta en los dos picos de producción, lo cual sugiere que en los protocolos para muestrear esta plaga y establecer algún programa de manejo en el cultivo del guayabo esta información debe considerarse a fin de no subestimar la presencia de la misma. Agradecimiento Se agradece al CONDES-LUZ por el financiamiento a través del Programa Museo de Artrópodos, No. CC-133.02. A la TSU Idelma Dorado por su colaboración en las labores de campo y laboratorio. Al Propietario de La Coruba, Sr. Esmeiro Villalobos, por permitirnos el uso de su plantación para esta investigación. Referencias
© 2005- Sociedad Venezolana de Entomología The following images related to this document are available:Photo images[em05007f3.jpg] [em05007f7.jpg] [em05007f2.jpg] [em05007f1.jpg] [em05007f4.jpg] [em05007f6.jpg] |
| |||||||||
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}