 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
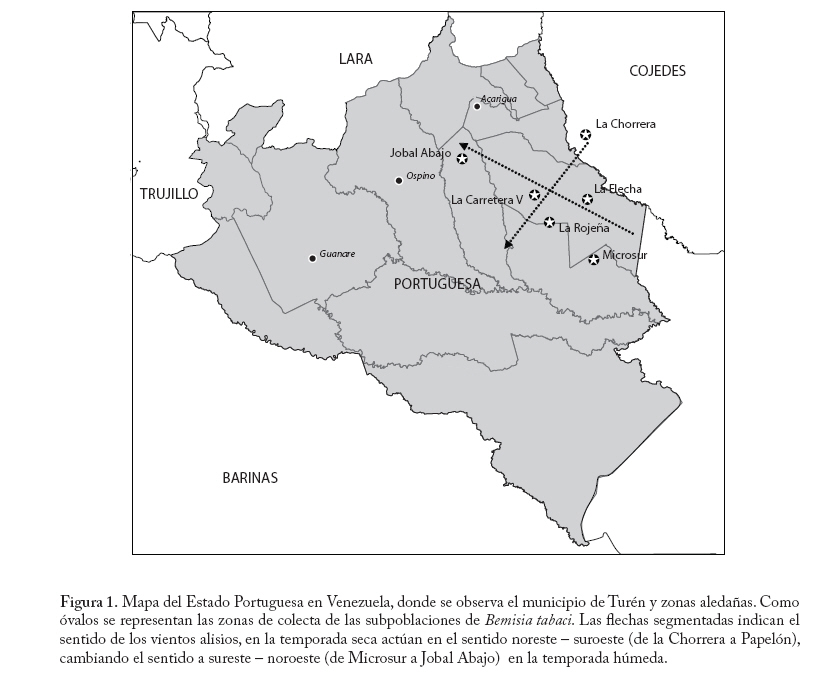
Entomotropica antes/formerly Boletín de Entomología Venezolana, Vol. 20, No. 3, December, 2005, pp. 249-263 La diversidad isoenzimática de Bemisia tabaci Gennadius 1889 (Hemiptera: Aleyrodidae) en el cultivo del ajonjolí (Sesamum indicum L) en Venezuela Isoenzimatic diversity of Bemisia tabaci Gennadius 1889 (Hemiptera: Aleyrodidae) on sesame plantations (Sesamum indicum L) in Venezuela. Luis Tovar1,2, Antonio Díaz2, Eustaquio Arnal3 y Catalina Ramis2 1Postgrado en Estadística, Fac. de Agronomía, Universidad Central de Venezuela. Apdo 4579. Maracay 2101, Aragua. Venezuela. luis_a_to@yahoo.com Code Number: em05027 Resumen Tovar L, Díaz A, Arnal E, Ramis C. 2005. La diversidad isoenzimática de Bemisia tabaci Gennadius 1889 (Hemiptera: Aleyrodidae) en el cultivo del ajonjolí (Sesamum indicum L) en Venezuela. Entomotropica 20(3): 249-263. Para cuantificar la diversidad y caracterizar la estructura genética de la población de Bemisia tabaci en la región productora de ajonjolí de Venezuela, se estudiaron siete subpoblaciones del insecto colectadas sobre el cultivo en diferentes localidades o ambientes. Se caracterizaron las frecuencias de los patrones (genotipos) de las isoenzimas a-esterasa, b-esterasa, enzima málica y malato deshidrogenasa, obtenidos en geles de poliacrilamida. La diversidad genética de toda la población (HT) fue de 0,42, entendida como la probabilidad de encontrar dos genotipos distintos al tomar aleatoriamente dos individuos de la población. La proporción de la diversidad genética total distribuida entre subpoblaciones fue de un 22%, indicando cierta fragmentación genética. Una prueba de independencia χ2 sirvió para detectar estadísticamente esta diferenciación, aunque no se observaron valores de H distintos entre localidades según la prueba de F (F=0,93; P=0,48). No se encontró correlación lineal entre la distancia genotípica de Hedrick (Dxy) y la distancia geográfica (Gxy) entre subpoblaciones según la prueba de Mantel, lo cual sugiere que la migración entre las localidades (distancias > 15 Km) podría tener menos efecto en la estructura genética de la población que los factores que se podrían considerar aleatorios. Por tanto, un modelo de aislamiento por distancia geográfica no puede ser sugerido. Posiblemente, factores que originan deriva genética aleatoria en cada localidad, entre ellos el efecto fundador, lluvias interanuales, quemas, control de malezas u otras actividades agrícolas, estarían condicionando la magnitud y estructura de la variación genética de la población de Bemisia tabaci en el cultivo del ajonjolí en Venezuela. Palabras clave adicionales: Aislamiento por distancia, Cojedes, diversidad genética, electroforesis, mosca blanca, Portuguesa, Turén. Summary Tovar L, Díaz A, Arnal E, Ramis C. 2005. Isoenzimatic diversity of Bemisia tabaci Gennadius 1889 (Hemiptera: Aleyrodidae) on sesame plantations (Sesamum indicum L) in Venezuela. Entomotropica 20(3): 249-263. To characterize the magnitude and structure of the genetic diversity existing within a Bemisia tabaci population in the sesame cultivated region, seven subpopulations of B. tabaci sampled on the crop in different localities or environments were studied. Patterns frequencies of α-esterase, β-esterase, malic enzyme and malate dehydrogenase systems were analyzed by polyacrilamide gel electrophoresis. The genetic diversity of the whole population (HT) was 0.42 taken as the probability of detecting two randomly chosen individuals with different genotypes. The proportion of total genetic diversity distributed among subpopulations was 22%, indicating some genetic fragmentation. A χ2 independence test was used to detect this differentiation, although different H values among localities were not detected by the F test (F=0.93; P=0.48). No linear correlation was found by the Mantel test between Hedrick genotypic distance (Dxy) and geographic distance (Gxy) among subpopulations, suggesting that migration among localities (distances >15 Km) could have less effect on population genetic structure than random factors. Therefore, the model of isolation by geographic distance cannot explain the phenomenon. Probably, factors that may generate random genetic drift, such as the founder effect, interannual precipitations, burnings, weed control and other agriculture activities are modifying the magnitude and structure of the genetic diversity of Bemisia tabaci population on sesame plantations in Venezuela. Additional Key words: Isolation by distance, Cojedes, electrophoresis, genetic diversity, Portuguesa, Turén, whitefly. Introducción En los últimos años, la mosca blanca Bemisia tabaci Gennadius (Hemiptera: Aleyrodidae) ha sido quizás la plaga que ha causado más pérdidas en la producción de cultivos a escala mundial (Salas y Arnal 2001; Brown et al. 1995). Bemisia tabaci presenta un complejo de razas con una estrecha y complicada relación (Brown et al. 1995; de Barro y Hart 2000), que se refleja en una alta variabilidad morfológica, biológica y genética, a tal punto que se ha generado la hipótesis de que es un género en proceso de especiación (Rosell et al. 1997). En Venezuela se ha planteado recientemente que Bemisia requiere revisión taxonómica (Arnal y Ramos 2000). Su hábito alimenticio es polífago y está adaptada a variados ecosistemas, por lo que puede estar asociada a hospederos anuales, bianuales o perennes (Arnal y Ramos 1997; Arnal et al. 1993a; Arnal et al. 1993b; Brown 1993). La adaptabilidad de este insecto a diferentes ambientes y la plasticidad de su ciclo de vida le ha permitido colonizar 506 hospederos distribuidos en 74 familias botánicas en todo el mundo. Se ha registrado en Venezuela en 112 malezas y 35 plantas cultivadas (Arnal y Ramos 1997; Arnal et al. 1993a; Arnal et al. 1993b), determinándose variaciónes morfológicas en la caja puparia, dependiendo del hospedero en que se haya criado (Arnal 1985). A inicios de la década de los noventa del siglo pasado, se reportó en Venezuela la aparición de grandes poblaciones de mosca blanca atacando cultivos de tomate (Edos. Falcón y Lara), que se supuso eran B. tabaci (Salas y Arnal 2001). Posteriormente, se comenzaron a observar crías masivas de Bemisia transmitiendo desórdenes virales y ocasionando daños severos en hospederos no tradicionales en el país. Desde entonces la producción del cultivo del ajonjolí (Sesamum indicum L.) se ha visto afectada por este insecto (Arnal et al. 1993b; Laurentin 1996). La región de producción de ajonjolí se encuentra en la localidad de Turén y zonas aledañas, estado Portuguesa, donde la mosca blanca incrementa sus poblaciones a mediados del mes de febrero (ciclo norte-verano). Los agricultores tratan de escapar del pico poblaciónal de febrero haciendo siembras tempranas entre octubre y noviembre (Pereira y Laurentin 2001). Sin embargo, enfrentan el riesgo de que lluvias extemporáneas dañen la germinación por encostramiento del terreno. Esta situación hace que el factor riesgo en el cultivo de ajonjolí aumente sensiblemente. Por otra parte, la aplicación de insecticidas es costosa y nociva para el medio ambiente, además de que pueden generarse individuos resistentes a los mismos (Dittrich et al. 1990). Entre las alternativas del manejo integrado de plagas, convendría estudiar los mecanismos de resistencia genética, donde Laurentin y Pereira (2002) han evaluado en Turén germoplasma mejorado de ajonjolí reportando posibles fuentes de resistencia. La cuantificación de la diversidad genética de la población de B. tabaci en Turén daría una idea de su potencial de adaptación al ambiente y a factores selectivos de origen antrópico. Una estimación de la diversidad, en especial, sería un indicador útil acerca de la capacidad del insecto para ‘vencer’ la resistencia que puedan ofrecer los cultivares de ajonjolí de la región o las líneas élites experimentales. Para el mejorador de plantas, esta información sugeriría la estrategia en cuanto al manejo de la resistencia parcial y/o completa que permita controlar la potencial aparición de nuevos biotipos del insecto, los cuales superen los genes ya conocidos como resistentes. De igual manera, la caracterización de la variabilidad genética en función de los elementos topográficos, ecológicos, ambientales, etc., constituye una herramienta para inferir procesos poblacionales que se crean estén influyendo o hayan influido en la diferenciación genética de organismos distribuidos en subpoblaciones geográficas (Bjørnstad et al. 1999; Koenig 1999). Por tales motivos, el objetivo del presente trabajo fue estimar la magnitud y estudiar la estructura de la diversidad genética de B. tabaci en el cultivo del ajonjolí en la región de Turén en Venezuela. Materiales y Métodos Zona de colecta La colecta se enmarcó entre los estados Portuguesa y Cojedes. En el estado Portuguesa se concentra el 90% de la superficie de ajonjolí sembrada en el país, específicamente en Turén y zonas aledañas (Figura 1). Con fines prácticos esta región ajonjolicera se denominará de ahora en adelante ‘Turén’. Turén presenta dos períodos climáticos distintos, uno de sequía entre noviembre y abril (norte-verano) y otro de lluvias (1 600 mm anuales, aproximadamente) entre mayo y octubre. Esta región no presenta accidentes geográficos y en la temporada de altas poblaciones de la mosca blanca los vientos alisios se presentan desde el nor-este hacia el sur-oeste. Además, en ella se realiza un laboreo agrícola intensivo, en el que el cultivo del ajonjolí se siembra en el período de sequía como un rubro de rotación a los de época de lluvias, principalmente maíz. Colecta del insecto La colecta de individuos se hizo al azar en cinco localidades alrededor de Turén (Edo. Portuguesa) y en una oportunidad se colecto en La Chorrera (Edo. Cojedes); en el período de sequía del año 2000, entre el 24 de febrero y el nueve de marzo (Cuadro 1, Figura 1). Todas las colectas se realizaron sobre el ajonjolí. En cinco localidades se realizó un solo muestreo, mientras que en La Carretera v se hicieron dos, espaciados por una semana. Los individuos colectados en cada localidad o ambiente diferente se denominaron ‘subpoblaciones’. Las siete subpoblaciones a su vez conformaron la población total de Turén (P-Turén). Cuadro 1. Datos de colecta de las subpoblaciones de Bemisia tabaci de la región Turén, Venezuela.
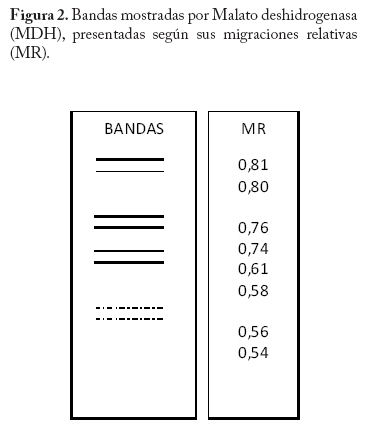
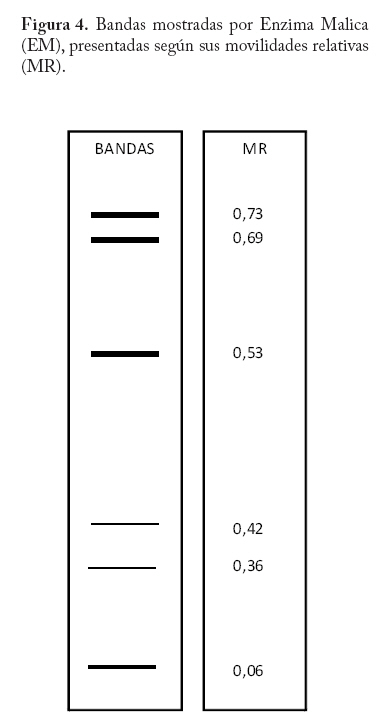
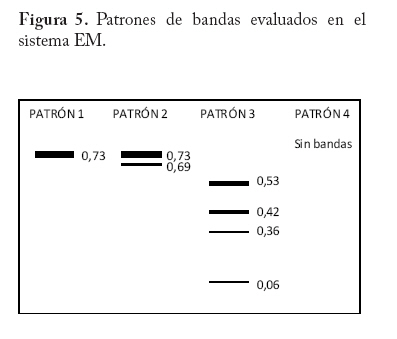
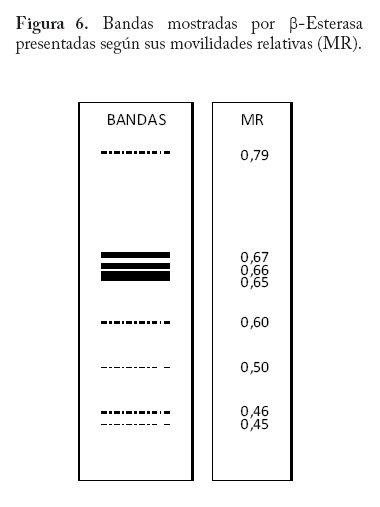
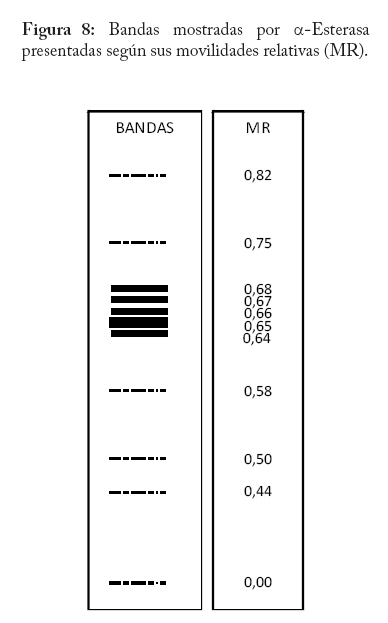
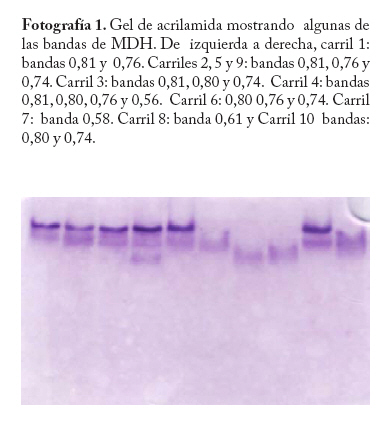
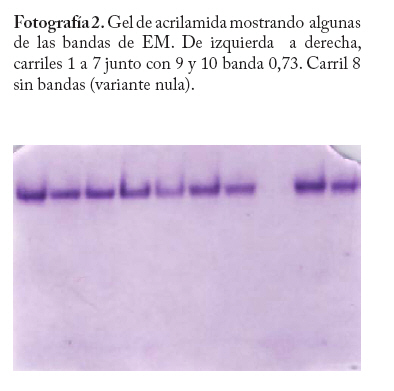
En campo se colectaron adultos directamente de la colonia a través de un aspirador de boca y se trasladaron al laboratorio a una temperatura menor o igual a 0 C° y guardados a -20°C hasta el momento del procesamiento de las muestras. En la localidad de Microsur, la colonia era en su mayoría ninfas, por lo que se trasladaron al invernadero para la obtención de adultos. Las cajas puparias de cada subpoblación se colectaron al mismo tiempo que los adultos y se enviaron al departamento de Protección Vegetal del Instituto Nacional de Investigaciones Agrícolas (INIA, Venezuela), donde se montaron e identificaron como Bemisia tabaci. La identificación la realizaron los entomólogos Eustaquio Arnal y Fidel Ramos sobre la base de la colección de moscas blancas de Venezuela. Electroforesis de isoenzimas Previo a la maceración de los adultos, las hembras se separaron de los machos con una lupa estereoscópica, diferenciándolas por las estructuras sexuales (clasper-aedeagus/ovipositor) y el grosor del abdomen. Con el fin de evitar posibles variaciónes de origen cromosómico (machos haploides), las hembras se colocaron individualmente en viales de 1ml de capacidad y fueron maceradas en 20 µl de solución tampón Tris-Borato-EDTA 0,1 M a pH 8 (Tovar 2000). Luego se añadió a cada vial 1 µl de Glicerol y 1 µl de Azul de Bromofenol y se centrifugó por 4 minutos a 3 000 rpm. Para la corrida electroforética se utilizó un sistema discontinuo en minigeles de poliacrilamida: 5% de poliacrilamida pH 6,8 en el gel superior y 7,5% a pH 8,8 en el gel inferior (Tovar 2000). Los minigeles se cargaron con todo el extracto disponible del vial. El tampón de los electrodos empleados fue Tris-glicina 1,5 M, pH 8,3 (Costa y Brown 1991). La corrida se realizó a 70 V constantes a 4 °C y detenida cuando el frente de migración alcanzaba el borde anódico del gel. Todo el proceso se realizó manteniendo una temperatura siempre menor a 4 °C. El revelado de los sistemas isoenzimáticos se hizo de la siguiente manera: Enzima málica (EM; EC: 1.1.140): 45 ml Tris 0,1 M, pH 8,0; 10 mg MTT; 8 mg NADP; 3 mg PMS; 1 ml MgCl2 10%; 10 mg ácido málico. Malato deshidrogenasa (MDH; EC: 1.1.1.37): 45 ml Tris 0,1 M, pH 8,0; 10 mg MTT; 10 mg NAD; 2 mg PMS; 1 ml MgCl2 10%; 10 mg ácido málico. α - esterasa (α-EST; EC: 3.1.1): 50 ml tampón fosfato 0,1 M, pH 6,3; 25 mg fast blue RR; 1 ml solución (0,75g α-naftil acetato, 30 ml acetona). β - esterasa (β-EST; EC: 3.1.1): 50 ml tampón fosfato 0,1 M, pH 6,3; 25 mg fast blue RR; 1 ml solución (0,75g β-naftil acetato, 30 ml acetona). Análisis Debido a que no se dispuso del control genético de los sistema enzimáticos, se hizo un registro cualitativo de la presencia versus la ausencia de cada patrón de bandas. Se trató a cada patrón como un genotipo, a la vez que no se incluyo en el análisis los patrones que no pudieron ser registrados objetivamente, debido a su pobre definición (ver Apéndice 1). Luego se estimó por conteo directo la frecuencia de cada genotipo en cada subpoblación por sistema enzimático y en P-Turén. Se analizaron entre 10 y 24 individuos por subpoblación y sistema enzimático, para un total de 542 en P-Turén. Se calcularon una serie de estimadores de la diversidad para cada sistema isoenzimático. El índice de diversidad genética usado para cada subpoblación cuantificó la probabilidad de encontrar un par de genotipos diferentes al azar, obtenido como h=1-∑pi2, en el que pi es la frecuencia del i-ésimo genotipo. Para caracterizar la estructura genética de la población P-Turén, la diversidad genética se descompuso en tres partes, la diversidad genética total (hT), y la contenida dentro (hD) y entre (hE) subpoblaciones. hD se calculó como el promedio aritmético de los estimados de h de todas las subpoblaciones por sistema enzimático; mientras que hE como la diferencia hT- hD. De igual manera, se calculó el cociente r(E/T)=hE/hT, que indicó la proporción de la diversidad total explicada por la diversidad entre subpoblaciones. Al promediar los valores de los distintos sistemas isoenzimáticos se generaron los estimados promedios de HT, HD, HE y R(E/T)=HE/HT en la población P-Turén; y por otra parte se obtuvo H para cada subpoblación. A su vez, se probó la heterogeneidad de las frecuencias genotípicas en las subpoblaciones mediante una prueba de independencia χ2, en la cual las subpoblaciones fueron confrontadas con las frecuencias de los distintos genotipos, en cada sistema isoenzimático. Para conocer acerca de la existencia de diferencias entre los valores de H entre subpoblaciones, se realizó un análisis de varianza de dos vías con los valores de h, en el que las subpoblaciones y los sistemas isoenzimáticos (bloques) fueron las fuentes de variación. Los cálculos se hicieron con el Proc ANOVA del programa SAS (Sas Institute 1988). Transformando el estimado de la diversidad genotípica HT en un estimado aproximado del índice de diversidad genética (D) de Nei, se podrían realizar comparaciones de interés a ser discutidas. El cálculo de D requiere de manera formal de la estimación previa de las frecuencias alélicas. Según Nei (1978), si se asume que cada sistema está codificado por un solo locus con dos alelos y la población se encuentra en equilibrio de Hardy y Weinberg, D se puede obtener mediante la relación D/HT = C = [2pq/(1-p4-4p2q2-q4)]. De tal manera que D = C*HT, donde p y q=1-p son las frecuencias alélicas. Se calculó la identidad genotípica de Hedrick entre todos los pares de subpoblaciones a través de la ecuación (ixy = ∑j Pj.x . Pj.y / 0,5 (∑j P2j.x + ∑j P2j.y), donde Pj es la frecuencia del j-ésimo genotipo en cada subpoblación (Hedrick 1971). Los valores de “ixy” se promediaron para todos los sistemas isoenzimáticos y se obtuvo Ixy. Esta identidad es el cociente entre la probabilidad de conseguir dos genotipos idénticos en dos subpoblaciones tomadas al azar y la probabilidad promedio de conseguir dos genotipos idénticos dentro de ellas. Por complemento se encontró Dxy, la distancia genotípica de Hedrick. La correlación lineal r entre Dxy y Gxy se obtuvo mediante la prueba de Mantel, utilizando los programas NTSYS (Rohlf 1990) y XL-STAT. Esta prueba calcula un producto z=Σni<j XijYij, en el que Xij y Yij son los elementos fuera de la diagonal de ambas matrices. Este valor es conocido como el producto Hadamard y al estandarizarlo por las respectivas desviaciones se obtiene el coeficiente de correlación lineal r entre las matrices (Sokal y Rohlf 1995). En el modelo de Mantel, las distancias genotípicas podrían ser explicadas por las geográficas mediante la siguiente ecuación Dxy = B0 + B1Gxy + E, donde los B`s son lo respectivos coeficientes de regresión y E representa el error aleatorio. r es usado como un estimador de B1. Por tanto, la hipótesis alternativa es el modelo de aislamiento por distancias, si se observa que a medida que aumentan las distancias geográficas disminuye la relación genética entre las subpoblaciones (Autocorrelación espacial). El rechazo de esta hipótesis ocurre cuando por encima del valor observado de r se presentan menos de un 5% de valores r generados por las permutaciones de las filas/columnas de la matriz que contiene a Dxy y Gxy. En este caso se emplearon 1.000 permutaciones. Resultados Diversidad Genética Las frecuencias genotípicas de las siete subpoblaciones o ambientes se muestran en el Cuadro 2. En el Apéndice 1 se muestra las bandas y los patrones registrados, así como fotografías de los geles. La población de B. tabaci analizada (P-Turén) fue polimórfica para todos los sistemas isoenzimáticos evaluados, con un promedio de cuatro genotipos por sistema. Ninguna subpoblación presentó todos los genotipos observados en la población. La Carretera v-1 y La Chorrera fueron las únicas subpoblaciones con cuatros sistemas polimórficos. Cuadro 2. Frecuencias de patrones isoenzimáticos en siete subpoblaciones de Bemisia tabaci de la región Turén, Venezuela.
(a) Los patrones fueron designados según números romanos. La población mostró una diversidad genética promedio (HT) de 0,42 (Cuadro 3). La diversidad genética promedio dentro de las subpoblaciones (HD) fue de 0,33 y la conseguida entre subpoblaciones (HE) de 0,09. Por lo tanto, el valor de RE/T fue igual a 0,22, señalando que de la variabilidad total de la población, un 22% se distribuyó entre las subpoblaciones y por complemento, cada subpoblación en promedio representó un 78% de la variabilidad de la población. Cuadro 3. Estimaciones de la diversidad genética para siete subpoblaciones de Bemisia tabaci colectadas sobre el cultivo del ajonjolí en la región Turén, Venezuela.
(a) Entre paréntesis el número de individuos utilizados. El intervalo de variación de H entre las subpoblaciones fue de 0,21-0,48 (Cuadro 3). Las subpoblaciones que mostraron estos valores extremos fueron colectadas en la misma localidad (La Carretera v), y en una semana de diferencia se observó la caída de H en el tiempo, asociada a la pérdida de algunos genotipos en β-EST y EM. Para los valores de H encontrados en este intervalo, el análisis de varianza no detectó diferencias significativas (F = 0,93; P = 0,50), es decir poco efecto de localidad y tiempo en la cantidad de variabilidad por subpoblación. La prueba de independencia (contingencia) entre las frecuencias genotípicas y las subpoblaciones arrojó resultados significativos en cada sistema (Cuadro 4). Esto indica que dichas frecuencias no fueron idénticas entre las subpoblaciones. De particular interés fue la comparación entre La Carretera v-1 y v-2, donde la prueba de contingencia detectó diferencias significativas en el sistema β-EST (χ2 = 10,39; P = 0,016), en el que se observó la pérdida de tres genotipos en una semana. Cuadro 4. Prueba de independencia o contingencia entre las frecuencias genotípicas y las subpoblaciones de la región Turén, Venezuela.
(1) Probabilidad asociada al valor χ2 Semejanzas genéticas La matriz con los valores promedios de la identidad genotípica de Hedrick (Ixy) y de las distancias geográficas (Gxy) se presentan en el Cuadro 5. El promedio de todas las comparaciones de Ixy fue igual a 0,82 y el intervalo de variación estuvo entre 0,60 (La Rojeña vs La Fecha) y 0,97 (La Chorrera vs Carretera v-2). El valor promedio de Ixy indica que un 82% de las comparaciones entre individuos de dos subpoblaciones distintas presentaron los mismos genotipos que los conseguidos en las comparaciones dentro de las subpoblaciones. Cuadro 5. Matriz de identidad genotípica de Hedrick (IXY) entre las subpoblaciones colectadas en la región Turén, Venezuela (bajo la diagonal) y las distancias geográficas en kilómetros (GXY) que las separan (sobre la diagonal).
Cuando se propuso a Gxy como una variación asociada a Dxy, no se detectó correlación lineal entre las mismas según la prueba de Mantel (Cuadro 6), lo que sugiere que, en la estructura poblacional de P-Turén la deriva aleatoria es un factor de mayor determinación que la migración entre las localidades. Se probaron varias combinaciones entre las subpoblaciones, en caso de que el fenómeno de aislamiento por distancia tenga mayor relevancia entre ciertas subpoblaciones en particular. En los gráficos de dispersión entre los Dxy y Gxy obtenidos en cada caso (no publicados), se observó el predominio del azar. Se puede destacar, que cuando se probo el modelo en poblaciones alienadas en un sentido perpendicular al de los vientos alisios (Figura 1 y Cuadro 6), donde los vientos se podrían sumar como barrera física a la distancia geográfica, la magnitud de los coeficientes de correlación r estimados, sus signos y valores de probabilidad P asociados no fueron adecuados para sugerir la hipótesis del aislamiento por distancia. Cuadro 6. Estimaciones del coeficiente de correlación r entre las distancias genéticas y las geográficas (autocorrelación espacial). El modelo de aislamiento de las poblaciones en la distancia no puede ser sugerido. Se muestran los valores de probabilidades asociados a las estimaciones P y en kilómetros (Km) se indica el intervalo de distancia geográfica.
Discusión Para el caso de P-Turén y sus subpoblaciones, se empleó el índice H para cuantificar la diversidad genotípica. El índice H estima la probabilidad de encontrar dos genotipos distintos en una localidad o subpoblación, de allí que el valor conseguido (HT = 0,42) sugiere que la población fue diversa. El complemento de HT (1-HT) mide el grado de predominancia de un genotipo en específico. Si se asume un equilibrio de Hardy y Weinberg, 1-HT mediría la predominancia de un homocigoto, lo cual está directamente relacionado con la posibilidad de perder variabilidad por deriva genética o selección. Bajo este supuesto, debido a que un 1-H de 0,58 de P-Turén está alejado del valor máximo de 1, se presenta una baja probabilidad de que los alelos de la población tiendan a perderse, si alguno de ellos lograra alcanzar una frecuencia cercana a la unidad. Con el fin de extender la comparación de la variabilidad genotípica estimada para B. tabaci en Turén a la de otros organismos, empleamos dos frecuencias contrastantes de p (0,5 y 0,95), para estimar una aproximación a D, el índice de diferenciación genética de Nei. Los valores obtenidos fueron 0,34 y 0,22, respectivamente; lo que sugiere que P-Turén presenta un D similar o mayor al de Drosophila y avispas (0,15), por encima del de mamíferos (0,037), reptiles (0,047) y aves (0,042) (Selander 1980). Dentro de la especie B. tabaci, P-Turén parece ser tan o más variable como un conjunto de 17 poblaciones de B. tabaci colectadas en diversas localidades de California y Florida, EE.UU (Perring et al. 1993). Las frecuencias alélicas de ocho loci polimórficos isoenzimáticos de seis poblaciones del biotipo A y 11 poblaciones del biotipo B fueron utilizadas para hacer el cálculo de D [valor no reportado por los autores]. El promedio por población del biotipo A fue igual a 0,216 y el del biotipo B de 0,103, para un promedio general de 0,143; valores que son similares o menores a los estimados de P-Turén (0,34 - 0,22). En este estudio, cada subpoblación representa el efecto del ambiente donde fue tomada la muestra sobre la variabilidad genotípica. Se cree que una notable variabilidad genotípica entre los sitios de colecta, podría ser un indicio de un proceso de diferenciación genética de la mosca blanca entre las regiones ajonjoliceras. Según la prueba de F, entre los valores promedios de H estimados en las subpoblaciones (0,21 - 0,48), cada ambiente presentó una variación genética similar a la promedio de la población P-Turén. Con la prueba de contingencia se detectaron diferencias significativas entre las frecuencias de los genotipos colectados en cada ambiente, lo cual indica diferenciación genética entre las subpoblaciones. El hecho de que la prueba de contingencia de las frecuencias genotípicas contraste con los resultados de la prueba de F entre los valores de H, muestra que hubo variabilidad en el orden relativo de los genotipos y las frecuencias de éstos variaron entre las subpoblaciones. Sin embargo, estas diferencias no se tradujeron en la predominancia de genotipo alguno que desviara los valores de H. Esto se explica, si se considera que diferentes arreglos o combinaciones de frecuencias de genotipos (incluso cualitativamente distintos entre sí) pueden generar valores similares de h. Los valores de RE/T como de χ2 sugieren que podrían existir variaciones originadas en los ambientes de colecta, lo que se traduce en una fragmentación geográfica de la diversidad. Las variaciones interlocales de la mosca blanca en la región ajonjolicera, representa alrededor de ¼ de la variación total y cuando se comparó esta tendencia con resultados de poblaciones de un mismo biotipo de B. tabaci colectadas en diferentes localidades, este patrón parece repetirse en poblaciones de España (Moya et al. 2001), Florida y California (Perring et al. 1993). Para los biotipos B y Q, las poblaciones de España arrojaron aproximadamente dos veces más variabilidad intra que interpoblacional. En las poblaciones de Florida y California, el componente de la diversidad genética total según el coeficiente de diferenciación genética GST (Nei 1973) fue de 13 y 21% dentro del biotipo A y B, respectivamente [valores obtenidos a partir de las frecuencias alélicas publicadas por Perring et al. 1993]. Adicionalmente, cuando la comparación entre las poblaciones arriba mencionadas se hizo entre diferentes biotipos (efecto biotipo), los resultados indican que esta fuente de variación podría ser un determinante de mayor peso para explicar la diferenciación que el efecto de las localidades o distribución geográfica. En España, se consiguió que independientemente de la localidad donde se colectaron las muestras, dos poblaciones pertenecientes a un mismo biotipo eran más similares genéticamente que dos poblaciones de biotipos distintos (Moya et al. 2001). De igual manera, el valor de GST aumentó a un 58% en las poblaciones de Norteamérica. Esto sugiere, que en B. tabaci la subdivisión de la población en subpoblaciones geográficas representa un efecto que origina una diferenciación genética no mayor al 25%, casos de Venezuela, España y EE.UU; mientras que esta diferenciación podría ser mayor el 50% cuando la variación observable es originada por el efecto biotipo, casos España y EE.UU (Perring et al. 1993; Moya et al. 2001). Con respecto a la dinámica de las frecuencias genotípicas, se tiene que las colectas realizadas con una semana de diferencia dentro del sembradío en La Carretera v, sugieren que la estructura genética no es estática en el tiempo. La prueba de independencia entre ambas subpoblaciones detectó diferencias significativas en β-EST, donde se perdieron tres genotipos. Mientras que en α-EST se perdió el genotipo más frecuente y a la vez apareció una variante de baja frecuencia, que limitó el cómputo del estadístico de prueba. Aúnque no se observaron diferencias entre los valores de H, se destaca que los estimados de estas dos subpoblaciones fueron los más contrastantes, relacionados negativamente en función del tiempo. Cuando se realizó la segúnda colecta, se observó el avance de la cosecha del cultivo en la región, lo que podría estar asociado la movimiento y/o perdida de insectos. En esta comparación particular observamos que, a pesar la estrecha relación entre estos dos ambientes, podrían existir en corto tiempo factores que condicionen diferencialmente las frecuencias genotipicas de la mosca blanca. Por otra parte, debemos tomar en cuenta que la diversidad genética estimada para la mosca blanca en la región de Turén, podría estar influenciada por el efecto del muestreo debido a la poca disposición de insectos (entre 10-24 para la estimación de h), al uso de individuos diferentes para promediar las repeticiones a causa de la limitante cantidad de proteínas obtenidas por individuo, entre otras. Los análisis de autocorrelación espacial mostrados en el cuadro 6 indican ausencia de correlación entre las distancias genéticas y las geográficas. Según Koenig (1999), la dispersión ha sido el factor más comúnmente asociado la teoría del aislamiento por distancia (modelo de piedras de paso). Por otro lado, el viento ha sido citado como un factor fuertemente asociado a la dispersión de la mosca blanca (Byrne et al. 1996). Se realizaron los cálculos usando varias arreglos entre las subpoblaciones, conociendo que el fenómeno de la autocorrelación espacial podría presentarse en mayor grado entre algunas localidades en especial. Sin embargo, en ningún caso se pudo suponer el modelo de aislamiento por distancia, incluso cuando los vientos alisios pudieron influir en el valor de r (Figura 1, Cuadro 6). Esto sugiere que, en el caso de P-Turén, el intercambio de individuos entre distancias de 64 a 15 kilómetros es de menor relevancia que la deriva aleatoria en cada localidad. Según Isaacs y Byrne (1998), la dispersión a largas distancias de B. tabaci parece ser de escasa importancia si se compara con las dispersiones a cortas distancias. Este comportamiento parece haber sido mantenido por la evolución, quizás como un mecanismo para colonizar nuevos hábitat sin causar una gran pérdida de individuos en la población (Isaacs y Byrne 1998). Si resulta cierta esta hipótesis, se requeriría más tiempo para que P-Turén encuentre un equilibrio entre la perdida por deriva y la ganancia por migración; ya que la falta de equilibrio detectada en este estudio, no implica que las poblaciones no estén en contacto. Otra posible causa de la falta de equilibrio entre la migración y la deriva podría ser la expansión del biotipo B de B. tabaci en los años 90 (Salas y Arnal 2001) en Venezuela, donde se cuenta al ajonjolí como un hospedero no tradicional colonizado por la mosca blanca luego de la aparición de este biotipo (Arnal et al. 1993b; Laurentin 1996). Igualmente, se requeriría más tiempo para que los procesos migratorios equilibren las perdidas por deriva. La deriva genética podría ser consecuencia de la disminución anual en el tamaño de la población, producto de las precipitaciones en la zona durante el período de mayo a agosto combinado con la siembra del maíz. La dinámica anual del clima en la región, conlleva a B. tabaci a enfrentar un proceso de extinción y recolonización del ambiente. El efecto de las precipitaciones es tan marcado en la región, que se han observado caídas drásticas en la densidad poblacional en época de sequía debido a lluvias extemporáneas (Linares 2002). Este efecto de deriva, también podría ocurrir de forma diferencial entre las localidades, ya que las lluvias en la región suelen ser cortas, intensas y localizadas. Además es necesario considerar, que las variaciones en la oferta alimenticia y/o otros disturbios antropocéntricos (quemas, inundaciones, control de malezas, plaguicidas, etc.) comunes en un área de alta intervención agrícola como Turén, pueden influir en las frecuencias genotípicas de forma muy particular en cada ambiente. Por tanto, El grado de fragmentación genética de P-Turén (22 %) podría explicarse, si se considera por ejemplo, el efecto de diversas inmigraciones con efectos fundadores distintos en cada localidad en conjunto con la diferenciación entre las localidades por deriva genética aleatoria. Sokal y Rohlf (1995) señalan que alrededor de siete subpoblaciones podrían producir el numero de permutaciones adecuadas para generar una distribución de los valores r que asuma en lo posible las propiedades de la distribución normal. En algunos casos se analizaron grupos pequeños de poblaciones en este estudio. Por otra parte, autores como Koenig (1999) y Catanéo et al (2003), discuten acerca de la poca disposición actual de datos que permitan el avance de la comprensión y por ende de los modelos que estudian la diferenciación genética espacial o la agregación espacial de fenómenos biológicos en general. En el ajonjolí se han detectado dos posibles variantes de resistencia a B. tabaci. Laurentin y Pereira (2002) y Laurentin y Pereira (2000) han conseguido evidencias de antibiosis y antixenosis. Esta combinación involucra una resistencia multifactorial generalmente más durable (van Endem 1987; de Ponti et al. 1990). Esta situación, es ventajosa para un programa de mejoramiento del ajonjolí, pues la alta diversidad genética estimada en la población P-Turén con una aparente no-estructura geográfica, sugiere la existencia de una población con un gran potencial para ‘vencer’ cualquier resistencia basada en uno o pocos genes de efecto mayor (van Endem 1987). No obstante, sería interesante conocer si la alta diversidad de P-Turén podría generar nuevos biotipos en contra de estos mecanismos de resistencia. Como consideración final, se plantea la necesidad de una serie de estudios sistemáticos en el tiempo y el espacio con el fin de generar más información acerca de los factores que condicionan la magnitud de la variación genética y la estructura de B. tabaci en Turén. Debemos tomar en cuenta que el diagnóstico entomológico de la fitoprotección contempla y valora la especie bajo estudio así como los organismos y ecosistemas que se relacionan a ella (hospederos alternativos, enemigos naturales, etc). Por ultimo, en poblaciones de España y EEUU el efecto biotipo ha sido de gran valor para explicar la distribución de la variabilidad genética, e incluso teniendo más peso que la ubicación geográfica de las poblaciones. Esta variable no fue abordada en este estudio. Para despejar estas incógnitas se plantea en un futuro estudiar con un mayor número de marcadores genómicos las variaciones aquí detectadas y correlacionar esta variación a la dinámica poblacional de B. tabaci en Turén. Agradecimientos Los autores desean expresar su agradecimiento a Kevin Gorman (ICAR, Rothamsted, U.K.) y Fidel Ramos (INIA-CENIAP,Venezuela) por sus amplias colaboraciones en el desarrollo de esta investigación. Igualmente agradecemos al profesor Hernan Laurentin (UCLA, Venezuela), por su ayuda en la colecta de los insectos y revisiones al manuscrito; así como a la profesora Aída Ortiz (UCV-FAGRO, Venezuela) e Ing.Agro. Ciro Hernández por sus colaboraciones en el estado Cojedes. Por último, se destaca la colaboración del Centro de Investigaciones en Biotecnología Agrícola (UCV-FAGRO) por disponer del presupuesto y las instalaciones para la ejecución de esta investigación. Figuras 2, 3, 4, 5, 6, 7, 8, 9 Fotografías 1, 2, 3, 4 Referencias
© 2005- Sociedad Venezolana de Entomología The following images related to this document are available:Photo images[em05027f6.jpg] [em05027f3.jpg] [em05027p1.jpg] [em05027f4.jpg] [em05027f5.jpg] [em05027p4.jpg] [em05027p3.jpg] [em05027f2.jpg] [em05027f7.jpg] [em05027f9.jpg] [em05027f8.jpg] [em05027f1.jpg] [em05027p2.jpg] | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}