 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Iranian Journal of Pharmacology & Therapeutics, Vol. 5, No. 1, 2006, pp. 1-4 Review Article Role of Cytokines in Pathophysiology of AsthmaShailaja Mahajan and Anita A. Mehta Current Author Address: Shailaja Mahajan, Department of Pharmacology, L.M. College of Pharmacy, Ahmedabad. Gujarat, INDIA. Received May 6, 2006; Revised May 29, 2006; Accepted June 5, 2006 Code Number: pt06001 ABSTRACT The worldwide incidence, morbidity and mortality of asthma are increasing dramatically. It is one of the most common disorders encountered in clinical medicine in both children and adults. It affects approximately 5% of the adult population in the western world and its reported incidence is increasing vigorously in many developed nations. A network of novel mediators known as ‘pleiotropic cytokines’ regulate the intermittent airway inflammation, bronchial smooth muscle hyperreactivity and bronchoconstricion underlying asthma. Preclinical and clinical data suggest that, the pathophysiological features of asthma are thought to result from the aberrant expansion of cytokines and chemokines. Cytokines are small extracellular signaling protein produced by different cells and possess a wide spectrum of biological activities due to their redundancy and pleiotropic properties. They are critical to the functioning of both innate and adaptive immune responses. Cytokines and their receptors exhibit very high affinity for each other. Because of this high affinity, picomolar concentrations of cytokines can mediate a biological effect. The major groups of cytokines are lymphokines, proinflammatory cytokine, inhibitory cytokines and growth factors. Pathophysiology of asthma is still poorly understood and its cause remains unknown. On this basis, this review will concentrate on the synthesis, release and functional role of each cytokine in the pathophysiology of asthma. Keywords: Asthma, Hyperreactivity, Bronchoconstriction, Cytokines Asthma is one of the few chronic diseases in the developed world that is increasing in prevalence, despite better understanding of its pathogenesis and improved treatments [1]. It is one of the most common disorders encountered in clinical medicine in both children and adults. It affects approximately 5% of the adult population in the western world and its reported incidence is increasing dramatically in many developed nations. The cost of the disease is substantial, and the market for the pharmaceutical industry is estimated at $5.5 billion/year [2-3]. Asthma is a very complex and difficult term to define in simple manner, currently it is considered to be a group of different disorders characterized by three major features: i) intermittent and reversible airway obstruction leading to recurrent episodes of wheezing, breathlessness, chest tightness and cough ii) bronchohyperresponsiveness (BHR) which is defined as an increased sensitivity to bronchoconstrictors such as histamine and cholinergic agonists and iii) airway inflammation [4]. Pathophysiology of asthma is still poorly understood and its cause remains unknown. Many scientific reports suggest that, it involves the activation of many inflammatory cells like mast cells, macrophages/monocytes, eosinophils, T-helper type-2 lymphocytes (Th2), dendritic cells, basophils, neutrophils and platelets. It is now increasingly recognized that structural cells may also be important sources of mediators in asthma. Airway epithelial cells, smooth muscle cells, endothelial cells and fibroblasts are all capable of synthesizing and releasing inflammatory mediators [5]. To date, greater than 50 different mediators have been implicated in asthma. It is now clear that cytokines play a critical role in orchestrating, perpetuating and amplifying the inflammatory response in asthma that is evident in the asthmatic airways [6]. It is considered as a novel mediator of chronic obstructive pulmonary disease (COPD), rheumatoid arthritis, inflammatory bowel disease and allergic inflammation [1]. Although insight into the pathophysiology of asthma has increased substantially over recent years, a number of issues remain to be further clarified. On this basis, our review will concentrate on a better understanding of the exact functional role of each cytokine in the sensitization process and in the complex relationship between inflammation, remodeling and altered airway behavior. This review also examines recent articles on the relationship of cytokines to allergy and asthma with particular emphasis on immune mechanisms involved in disease development in early life. CytokinesCytokines are small, extracellular signaling proteins usually less than 80 KD in size and many are glycosylated. These mediators play an important role in the co-ordination and persistence of inflammation in asthma, although the precise role of each cytokine remains to be determined [8]. Cytokines are regulatory peptides so can be produced by virtually every nucleated cell type in the body and they have pleiotropic regulatory effects on hematopoietic and many other cell types that participate in host defense and repair processes [9]. Cytokines often possess overlapping biological activities, exert different effects at different concentrations, can synergize or antagonize the effects of other cytokines and regulated in a complex manner and function via cytokine cascade [10]. Cytokines and their receptors exhibit very high affinity for each other. Because of this high affinity, picomolar concentrations of cytokines can mediate a spectrum of biological effect. They affect closely adjacent cells, and therefore function in a predominantly paracrine fashion. They may also act at a distance (endocrine) and have effects on the cell of origin (autocrine) [11]. The major groups of cytokines are-Lymphokines, Proinflammatory cytokines, Inhibitory cytokines, Growth factors and Chemokines. A. Lymphokines or Type-2 T-Helper Cell Derived CytokinesLymphokines are cytokines that are produced by T-lymphocytes, although it is now recognized that many other cell types may release these cytokines. They play an important role in immunoregulation. Kips [12] reported that within the range of cytokines produced by Th2, IL-4 and IL-5 have received considerable interest to date. Other cytokines from this category includes IL-2, IL-3, IL-4, IL-5, IL-7, IL-9, IL- 15, IL-16 and IL-17. Interleukin-2 Synthesis and release. IL-2 is the short chain a-helical bundle with a size of 4-17 KD. It is produced and secreted primarily by mature T-cells particularly Th0 and Th1 cells [13]. It is considered as a primary growth factor and a potent modulator of T-cell and natural killer (NK) cell function, and plays a major role in the immune response. IL-2 can be synthesized by eosinophils and by airway epithelial cells [14]. In vitro, IL-2 is secreted by B-lymphocytes under certain conditions. IL-2 is also secreted by antigen-activated T cells 4 to 12 h after activation, accompanied later by up-regulation of high affinity IL-2 receptors on the same cells. Interaction between two IL-2 receptors induces proliferation of T cells, secretion of cytokines, and enhanced expression of receptors for other growth factors, such as insulin. Release of excess IL-2 leads to proliferation of T (including NK) cells, B-lymphocytes, differentiation of monocytes/macrophages as well as stimulation of IL-1 secretion [13, 15]. Role in asthma.IL-2 levels are increased in bronchoalveolar lavage fluid from patients with symptomatic asthma [16]. Robinson et al [17] reported that, increased bronchoalveolar lavage cells expression shows presence of IL-2 mRNA. It has been also reported that high levels of IL-2 and IL-4 mRNA-positive bronchoalveolar lavage cells are observed in steroid-resistant asthmatics, compared with steroid-sensitive asthmatics [18]. A significant increase in IL-2 mRNA positive cells is observed in asthmatics after allergen challenge [19]. Interleukin-3Synthesis and release.IL-3 is a short chain a-helical bundle having molecular mass 20-26KD. T-cells, NK cells and mast cells are major sources of IL-3. But the prominent source of IL-3 appears to be activated T-cells although other cell types have also been reported to produce it [20]. Role in asthma. IL-3 stimulates the formation of mixed colonies of neutrophilic granulocytes, macrophages, megakaryocytes and erythrocytes. IL-3 synergizes with GCSF, IL-1 and IL-6 to stimulate primitive cells. An increase in the number of cells expressing IL-3 mRNA has been reported in mucosal biopsies and in bronchoalveolar lavage cells from patients with asthma [21]. Interleukin-4Synthesis and release.IL-4 is a short chain a-helical bundle having molecular weight 18 KD. The major cellular sources of IL-4 are thymocytes, mature T-cells, mast cells, basophils and CD4+ Th2 cells. IL-4 is thought to be an upstream cytokine that regulates allergic inflammation by promoting Th2 cell differentiation and IgE synthesis [22]. IL-4 synthesis can be induced by stimulation of the antigen receptors on T-lymphocytes and by IgEFc receptor cross linking in mast cells and basophils. IL-4 was originally identified as a B-cell growth factor, which drives the optimal stimulation of B-cells by antigen [23]. Interestingly; corticosteroids enhance the capacity to induce IL-4 synthesis from CD4+ T-cells [24]. In addition, IL-4 stimulates the expression by B cells of major histocompatibity complex (MHC) Class-II molecules, B7, CD40, surface IgM and low affinity receptor, resulting in enhance antigen presenting capacity of B-cells [25].
Role in asthma.IL-4 demonstrates a broad range of biological activities, in general terms; it can be described as the main cytokine involved in the pathogenesis of allergic disorders [26]. Additional effects that seem of particular importance for asthma include stimulation of mucus producing cells and fibroblast, thus also implicating IL-4 in the pathogenesis of airway remodeling [27]. Another potentially important activity of IL-4 in allergic inflammation is its ability to induce the expression of vascular cell adhesion molecule-1 on endothelial cells. This will produced enhanced adhesiveness of the endothelium for T-cells, eosinophils, basophils and monocytes, which are characteristics of allergic reactions [28].Because of these propertiesIL-4 has long been considered as a potential target in allergies and asthma, numerous in vivo studies highlighted its role in IgE production [29]. However, it soon became clear that the role of IL-4 in IgE production could not be extended to airway hyperreactivity, which remains the hallmark of asthma. Over expression of IL-4 in lungs leads to a lymphocytic and eosinophilic inflammation, but without airway hyperreactivity [30]. Shi et al. [31] reported that, inhalation of recombinant human IL-4 induced airway eosinophilia and caused some degree of bronchial hyperresponsiveness in atopic asthmatics. Thus, an essential effector role for IL-4 in experimental asthma has been difficult to show, although IL-4 appears to play an important role in TH2 cell development and recruitment to the airways [32]. Interleukin-5Synthesis and release.IL-5 is the antiparallel dimmers having molecular weight 45-50KD. Activated T-helper cell populations are the main source of IL-5, but secretion from other cell types have been reported like eosinophils, mast cells, CD4+ and CD8+ T-cells [12]. However, IL-5 is crucial in regulating the eosinophilic response both in vitro and in vivo, as seen during helminthes infections. IL-5 induces activation and survival of eosinophils and marked proliferation of eosinophil precursors [33]. Transgenic mice in which IL-5 is constitutively expressed in all T cells show a profound and lifelong eosinophilia, with large number of eosinophils in the blood, spleen, and bone marrow. IL-5 is produced by T lymphocytes; in asthmatic airways and increased expression of IL-5 mRNA has been demonstrated in CD41 T cells, using in situ hybridization [34]. Bronchoalveolar lavage CD41 and CD81 T cells can also secrete IL-5 [35]. Role in asthma.IL-5 hasbeen shown to increasethe percentage of eosinophils in sputum and augment airway hyperresponsiveness in asthma [31, 36]. IL-5 is a cytokine that is highly specific for eosinophilic inflammation and antibodies that block IL-5 actions are effective in reducing eosinophilic inflammation and airway hyperresponsiveness (AHR) in various species [37]. Recently, studies of a humanized anti-IL-5 monoclonal antibody (mepomizulab) in asthmatic patients have confirmed its extraordinary efficacy in reducing eosinophils in the circulation and the airways, with surprisingly no reduction in response to allergen or in AHR [38]. Collin et al. [39] found that IL-5 may play an important role in eosinophil maturation, chemoattraction and activation in asthma. It may underlie bronchial hyperreactivity and also interact with other eosinophil chemoattractants and activators, such as chemokines, to activate and induce chemoattraction of eosinophils. Studies of transgenic mice expressing IL-5 from lung epithelial cells showed elevated levels of IL-5 in bronchoalveolar lavage fluid and serum, lung histopathological changes reminiscent of asthma, and base-line airway hyperresponsiveness [40]. Elevated IL-5 concentrations have been reported in bronchoalveolar lavage fluid from symptomatic but not asymptomatic asthmatics [41]. Long-term clinical studies are now in progress and should clarify the therapeutic benefit of IL-5 antagonism in asthma. Interleukin-7Synthesis and release.IL-7 is a cytokine produced by stromal cells of the thymus, bone marrow and keratin is known to play a key role in T-cell development. IL-7 production by human stromal cells is induced by IL-1 and by tumor necrosis factor alpha, which has been found to enhance HIV production in vitro. IL-7 induced T-cell proliferation and up-regulated CXCR4 expression in peripheral blood mononuclear cells in vitro [42].
Role in asthma. There are no data specific to asthma. Interleukin-9Synthesis and release.IL-9 belongs to the family of 4-helix bundle cytokines having molecular mass of 14 KD. A variety of cell types including T-cells, mast cells, eosinophils and neutrophils can produce IL-9. However, the major cell source of IL-9 is CD4+ T-lymphocytes [41]. Strong expression of IL-9 can be observed during stimulation of human peripheral blood mononuclear cells with antiCD3 antibodies. It could be further enhanced by the presence of various T-cell mitogens [43]. Role in asthma.IL-9 plays an important role in airway hyperresponsiveness, mucus hypersecretion and airway remodeling [44]. The chromosomal region 5q31-33 has been identified to contain candidate genes for AHR [45]. This region contains several gene candidates for cytokines, growth factors and growth factor receptor that may play a crucial role in asthmatic inflammation [46]. The IL-9 gene has been localized to this region [47]. Several reports have demonstrated upregulation of mucus production by IL-9 [48]. Recently, the mechanism by which IL-9 leads to mucus over production has been further elucidated. Expression of the calcium- activated chloride channel mCLCA3 in the lungs of a murine model of allergic asthma was associated with AHR and mucus overproduction. In IL-9 transgenic mice induction of mCLCA3 has been identified in the lung epithelium [49]. Increased expression of IL-9 supports eosinophil infiltration and survival [50]. IL-9 induces the release of T-cell chemoattractant factor from bronchial epithelial cells thus contributing to inflammation [51]. Interleukin-13Synthesis and release. IL-13 is produced by activated T-lymphocytes, B-lymphocytes and mast cells. In the mouse, almost exclusively Th2 clones express IL-13, however, in humans it can be expressed in both Th1 and Th2 lymphocyte clones [52]. Role in asthma.IL-13 is a cytokine closely related to IL-4 that binds to IL-4Ra receptors and is also expresses by Th2 cells from asthma patients [53]. IL-13 has a variety of pro as well as anti-inflammatory effects. IL-13 is present in increased amounts in asthmatic airways and has very similar biological activities to IL-4. Increased expression of IL-13 mRNA has been reported in the airway mucosa of patients with atopic and nonatopic asthma [54, 55]. In addition, levels of IL-13 together with IL-4 are increased after segmental allergen challenge of patients with asthma [56]. There is a significant correlation between eosinophil counts and levels of IL-13. In allergen induced airway changes, it has been hypothesized that IL-4 is crucial for the initial Th2 development during primary sensitization but IL-13 release might prove more important during secondary antigen exposure [57]. De Vries [58] found that IL-13 promotes goblet cell hyperplasia and mucus glycoprotein accumulation in the airways after exogenous administration in mice or increased endogenous production in IL-13 transgenic rats. Interleukin-15Synthesis and release.IL-15 is a pleiotropic cytokines that promotes activation and proliferation of and cytokines release from various subsets of T-cells, NK-cells, mast cells and B-cells [59]. Role in asthma.IL-15 is reported to be synergistic with IL-12 to induce proliferation of murine Th1 clones. Aberrant IL-15 expression was observed in patients with inflammatory autoimmune disease such as rheumatoid arthritis and inflammatory bowel disease. Ryotaro et al. [59] reported that overexpression of IL-15 in vivo suppresses Th2-mediated-allergic airway response via induction of CD8+ T cell mediated Tc1 response. Interleukin-16Synthesis and release. Previously IL-16 was known as lymphocyte chemoattractant factor. It was first identified as a product of peripheral blood mononuclear cells following mitogen and histamine stimulation in vitro [60]. Subsequently; it was shown to be produced by CD8+ T cells following stimulation with histamine and serotonin in vitro. IL-16 can also be produced by epithelial cells [61], eosinophils and mast cells [62]. Role in asthma.Elevated levels of IL-16 have been found in BAL fluid obtained from asthmatic subjects following allergen and histamine challenge [63]. In stable atopic asthmatic subjects there is predominant expression of IL-16 mRNA and immunoreactivity in airway epithelium [64]. In the ovalbumin sensitized and exposed mouse model IL-16 immunoreactivity was detected in the airway epithelium and an anti-IL-16 antibody prevented OVA specific IgE responses, bronchial hyperresponsiveness, but not airway eosinophilia [65]. Interleukin-17Synthesis and release.To date, no scientific report suggests the involvement of IL-17 in asthma pathophysiology; however some studies have shown its role in the synthesis of cytokines relevant to asthma. IL-17 is a CD4+ T cell derived cytokine which stimulates NF-kB and IL-6 production in fibroblasts and co-stimulates T cell proliferation [66]. It stimulates epithelial, endothelial, and fibroblastic cells to secrete cytokines such as IL-6, IL-8, GM-CSF, and PGE2 [67]. IL-17 increases the release of NO in cartilage from patients with osteoarthritis via NF-kB activation. In the presence of IL-17, fibroblasts can sustain the proliferation of CD34+ haematopoietic progenitors and their preferential maturation into neutrophils [68].
B. Proinflammatory Cytokines Another group of cytokines that needs to be considered are the proinflammatory cytokines such as IL-1, TNF - α , IL-6, IL-11, GM-CSF and SCF. They may be important in disease severity and resistance to anti-inflammatory therapy in asthma. The pleiotropic activities of these cytokines include proinflammatory effects such as leukocyte recruitment through upregulation of adhesion molecules on vascular endothelial cells and induction of cytokine and chemokine synthesis [69]. Interleukin-1Synthesis and release.There are two distinct forms of IL-1 (a andb).The structure of each IL-1 is b-bared shaped with 17KD molecular weight. The major cellular sources of IL-1 are monocytes, macrophages, neutrophils, eosinophils, mast cells, platelets, lymphocytes, NK cells, endothelial cells, airway smooth muscle cells and vascular smooth muscle cells [8]. These two subtypes of IL-1 are derived from two different genes. Monocytes produced 10 times as much IL-1b as IL-1a [70]. Air pollutants like nitrogen dioxide can stimulate epithelial cells express IL-1b while eosinophils can produce IL-1a [71]. A wide variety of stimuli including IL-1 itself [72] TNFa [73], GM-CSF, endotoxin and phagocytosis can increase the expression of IL-1 in monocytes/macrophages [72, 73]. Role in asthma.Bhandari [74] reported the role of IL-1 in hyperoxic lung injury in animal models. Hyperoxic (95% O2) exposure of alveolar macrophages obtained from adult monkeys via bronchoalveolar lavage for 24 hr resulted in significantly increased release of IL-1b. Patients with symptomatic asthma show increased level of IL-1b in BAL fluid compared with patients with asymptomatic asthma [75]. Increased expression of IL-1b in asthmatic airway epithelium has been reported, together with an increased number of macrophages expressing IL-1b [76]. Also IL-1b induces airway neutrophilia and increase airway responsiveness selectively to bradykinin in the rat [77]. Tumor Necrosis Factor-a (TNF - α)Synthesis and release.Two major forms of TNF are TNF - α and TNF-b, which have only 35% amino acid homology but bind to similar receptors. TNF - α was previously known as cachectin. It is expressed as a type II membrane protein attached by a signal anchor transmembrane domain in the propeptide [78]. TNF - α is produced by many cells including macrophages, T lymphocytes, mast cells, and epithelial cells, but the principal source is the macrophage. TNF - α is released from cells by proteolytic cleavage of the membrane bound form by a metalloproteinase, TNF - α converting enzyme (TACE). Inactivation of the TACE gene compromises the ability of cells to produce soluble TNF - α [79].The secretion of TNF - α by monocytes/macrophages is greatly enhanced by other cytokines such as IL-1, GM-CSF and IFN-g. Ohkawara et al. [80] showed that human eosinophils are also capable of releasing TNF - α, together with airway epithelial cells. Role in asthma.TNF - α being a proinflammatory cytokine is present abundantly in asthmatic airways. There is evidence that IgE triggering in sensitized lungs leads to increased expression in epithelial cells in both rat and human [81]. TNFa is present in the BAL fluid of asthmatic patients [82]. It is reported that TNFa is also released from alveolar macrophages of asthmatic patients after allergen challenge [83]. Furthermore, both monocytes and alveolar macrophages show increased gene expression of TNFa after IgE triggering in vitro that is enhanced by INF-γ. In vitro studies also indicate that TNF - α plays a role in bronchial hyperresponsiveness and airway remodeling in asthma [84]. Interleukin-6Synthesis and release.All IL-6 type cytokines are polypeptides with molecular masses of about 20 kD. It is secreted by monocytes/ macrophages, T cells, B cells, fibroblasts, bone marrow stromal cells, keratinocytes, and endothelial cells [74]. Epithelial cells also appear to produce IL-6 [85]. Human airway smooth muscle cells, upon activation with IL-1b or TGF-β, can release IL-6 [86]. Major basic protein secreted from eosinophils can interact with IL-1 or TGF to increase IL-6 release from fibroblasts [87]. IgE-dependent triggering stimulates the secretion of IL-6 from both blood monocytes and alveolar macrophages in vitro [88]. Role in asthma.IL-6 was originally described for its antiviral activity, its effects on hepatocytes, and its growth-promoting effects on B-lymphocytes and plasmacytomas. Now it has been accepted that IL-6 is released from alveolar macrophages from asthmatic patients after allergen challenge [89] and increased basal release, compared with nonasthmatic subjects [90]. Increased levels of IL-6 can be measured in nasal washings from children after rhinovirus infection. In addition, IL-6 mRNA expression and an increase in NFkB DNA-binding activity can be induced by rhinovirus infection of cells in vitro [91]. Interleukin-11Synthesis and release.The structure of IL-11 is 4a-helical with 19-23kD molecular mass. IL-11, which is distantly related to IL-6, is produced by fibroblasts, epithelial cells and human airway smooth muscle cells when stimulated by IL-1 and TGF-β1 [92]. In vitro studies have shown that IL-11 gene is expressed in a variety of cells of mesenchymal origin. Furthermore, IL-11 gene expression can be induced and/or unregulated by several inflammatory cytokines and agonists as well as hormones [93].
Role in asthma.IL-11 can be detected in BAL fluid during upper respiratory viral infections in humans and induces non-specific bronchial hyperresponsiveness in mice [94]. Tang et al. [95] reported that targeted expression of IL-11 in mouse airways leads to a T cell inflammatory response with airway remodeling, local accumulation of myofibroblasts, and airways obstruction. Adult transgenic mice that overexpress IL-11 in lungs were found to have significantly better survival in hyperoxia [96]. Granulocyte Monocyte Colony Stimulating Factor (GMCSF)Synthesis and release.GMCSF is a pleiotropic cytokine that can stimulate and regulate growth, proliferation, maturation and function of hematopoietic cells.Several airway cells produce GMCSF, including macrophages, eosinophils, T-lymphocytes, fibroblast, endothelial cells, airway smooth muscle cell and epithelial cells [97]. Role in asthma. Davieset al. [1] suggested that GM-CSF and IL-8 influence both eosinophils chemoattraction and activation in vivo and participated in the etiology of bronchial hyperresponsiveness in mild asthma. Sousa et al. [98] found increased expression of GM-CSF in the epithelium in bronchial biopsies from asthmatic patients and in T lymphocytes and eosinophils after endobronchial challenge with allergen [99]. Increased circulating concentrations of GM-CSF have been detected in patients with acute severe asthma [100], and peripheral blood monocytes from asthmatic patients secrete increased amounts of GM-CSF [101]. In addition to its release in asthmatic airways, GM-CSF can be demonstrated to have various effects in asthma. GM-CSF also has LTC4-enhancing activity for eosinophils in the supernatant of cultured asthmatic alveolar macrophages [102]. Stem cell factor (SCF)Synthesis and release. SCF previously known as c-Kit ligand is produced by bone marrow stromal cells, fibroblasts and epithelial cells, such as nasal polyp epithelial cells [103]. Role in asthma.There is very little information on the expression of SCF in asthmatic airways. SCF is expressed in the epithelium of nasal polyps removed from patients with allergic rhinitis [104]. C. Inhibitory CytokinesWhen evaluating the functional role of cytokines in asthma, it is important to bear in mind that they act within a network of mutually interacting cytokines. Allergic airway inflammation might, therefore, be induced by not only increased expression of Th2 cytokines but also decreased expression of counter acting ones. Interesting cytokines in this respect include IL-10, IL-12, IL-18 and IFN-g. Interleukin-10Synthesis and release.IL-10 is long chain-a helical structure having molecular weight 34-40kD. In humans Th0, Th1, and Th2-like CD4+ T cell clones, cytotoxic T cells, activated monocytes and peripheral blood T cells including CD4+ and CD8+ T cells have the capacity to produce IL-10 [105]. IL-10, previously known as cytokine synthesis inhibitor factor (CSIF) was originally identified as a product of murine T helper (Th2) clones that suppressed the production of cytokines by Th1 clones responding to stimulation of antigen [106]. Mast cells also have the capacity to produce IL-10. Constitutive IL-10 secretion occurs in the healthy lung with the major source being the alveolar macrophage; however, the circulating monocyte elaborates more IL-10 than the alveolar macrophage [107]. Role in asthma.IL-10 is a pleotropic cytokine that has the potential to downregulate both Th1 and Th2-driven inflammatory processes [108]. It is of interest that IL-10 might also have a beneficial effect on airway remodeling, as it has been shown to reduce collagen type-I synthesis and vascular smooth muscle proliferation [109]. The precise functional role of IL-10; however appears to be somewhat unclear. Whether IL-10 expression is changed in asthma is uncertain, as in some studies reduced and in others increased bronchoalveolar lavage fluid levels were found [110]. IL-10, administered exogenously at the time of secondary antigen presentation, reduces antigen-induced airway eosinophilia in animal models [111], where as airway eosinophilia and total serum IgE levels are increased in sensitized IL-10 knockout mice [112]. However, the effects of IL-10 on airway hyperresponsiveness are somewhat more contradictory. In some studies, endogenous production of IL-10 was shown to dampen airway responsiveness [113] whereas others have shown that IL-10 enhances the allergen induced increase in airway responsiveness, despite the reduction in eosinophil recruitment [114].
Interferon-γ (INF-γ) Synthesis and release. IFN-g was originally identified as a product of mitogen-stimulated T lymphocytes that inhibited viral replication in fibroblasts. The only known sources of IFN-g are CD4+ and CD8+ T cells and natural killer cells [5]. Role in asthma. In asthmatic patients nebulized IFN-g reduces the number of eosinophils in BAL fluid indicating its therapeutic potential in asthma [115]. The reduced production of IFN-g has been also reported in asthmatics [116]. This suggests that defective IFN-g production may be important in asthma [117] although no polymorphisms of the IFN-g gene have been associated with asthma [118]. Administration of exogenous IFN-g prevents airway eosinophilia and hyperresponsiveness after allergen exposure in mice [119]. Interleukin 12Synthesis and release.IL-12 plays an important role in Th1/Th2 differentiation during primary allergen presentation [12]. IL-12 is secreted by antigen presenting cells including-lymphocytes, monocytes/ macrophages, and dendritic cells [120]. It has been also reported that IL-12 is an inducer of IFN-g synthesis by resting human peripheral blood mononuclear cells in vitro. The major antigen presenting cell involved in the process of sensitization to aeroallergens is the dendritic cells present in the airway epithelium. Mucosal dendritic cells demonstrate high antigen uptake but low antigen presenting capacity [12]. Role in asthma.IL-12 expression has been shown to be reduced in bronchial biopsy sample from asthmatic individuals. It is also reported that, IL-12 inhibited IgE synthesis [121]. In vivo animal models have confirmed that exogenous administration of IL-12 during the primary sensitization process, suppresses allergen induced Th2 development, favoring Th1 differentiation [122]. Interestingly however, even when administered during secondary antigen exposure, IL-12 retains the capacity to inhibit allergen-induced eosinophilia and airway hyperresponsiveness [123]. These effects of IL-12 are largely mediated by IFN-g. In mice IL-12 administered at the time of allergic sensitization decreased specific IgE, tracheal ring responsiveness to acetylcholine, and eosinophilia in BAL fluid. In recently reported antigen-challenge study in mild asthma, IL-12 was given systematically in incremental doses, as advocated to reduce toxicity. Results showed a substantial reduction in circulatory blood eosinophil count and also IL-12 did not influence the antigen induced early or late asthmatic response [124]. Interleukin-18Synthesis and release.IL-18 is secreted by macrophages and was initially describedas INF-g releasing factor [125]. IL-18 (IFN-g inducing factor) is a cytokine that is a potent inducer of IFN-g production and plays an important role in Th1 responses [126]. Human IL-18 has been cloned from normal human liver cDNA libraries using murine IL-18 cDNA clones. IL-18 is synthesized as a precursor molecule without a signal peptide and requires the IL-1-converting enzyme (caspase-1) for cleavage into a mature peptide. Recombinant human IL-18 induces IFN-g production by mitogen-stimulated peripheral blood mononuclear cells, enhances natural killer cell cytotoxicity, increases GM-CSF production, and decreases IL-10 production. IL-18 also activates NF-kB in murine Th1 cells for enhancement of IL-2 gene expression by Th1 cells [133]. IL-18, together with IL-12, induce anti-CD40-activated B cells to produce IFN-g, which inhibits IL-4-dependent IgE production [127]. Role in asthma.Sanjive showed [128] the effect of IL-18 on suppressing eosinophil recruitment in the mouse model of asthma. However, evidence suggests that IL-18 augments eosinophil recruitment. Zhang and Liu [129] found that IL-18 is high in perennial allergic rhinitis patients and elevated levels of IL-18 suggest its role in the expression of persistence and exacerbation of allergic inflammation. It has been also suggested that IL-18 may play a potential role to activate immunologic responses and may reflect disease activity in mild and moderate asthma exacerbation [130]. D. Growth FactorsGrowth factors are involved in proliferation and differentiation of smooth muscle cells originating from a variety of tissues, including the vasculature and the airways [131]. They are potential contributors to the increased airway smooth muscle (ASM) mass, as found in patients suffering from persistent severe asthma, by stimulating airway smooth muscle proliferation [132]. The different growth factors involved in the pathophysiology of asthma are, Platelet derived growth factor, Transforming growth factor, Nerve growth factor, Fibroblast growth factor, epidermal growth factor and insulin like growth factor. Platelet-derived growth factor (PDGF)Synthesis and release. The sources of PDGF include platelets, macrophages, endothelial cells, fibroblasts, airway epithelial cells and vascular smooth muscle cells [133]. PDGF is released from many different cells in the airways and consists of two peptide chains, so that different cells may secrete AA, BB, or AB dimers. Various stimuli, such as IFN-γ from alveolar macrophages, hypoxia, basic FGF (bFGF), and mechanical stress for endothelial cells, and serum, TNF - α, IL-1, and TGF-β for fibroblasts can induce PDGF release [134]. Role in asthma.Levels of PDGF-AA, -AB, and –BB are not increased in asthma, and immunohistochemical analysis of PDGF-AA and -BB and PDGF receptor and b-subunits does not reveal increased expression [135]. A potential source of PDGF B chain has been identified as eosinophils in bronchial biopsies from patients with asthma [136]. This, suggest their ability to express TGF-β, has raised the possibility that eosinophils are involved in the airway remodeling of asthma. Transforming growth factor-β (TGF-β)Synthesis and release. Eosinophils, neutrophils, airway smooth muscle cells, lung fibroblast, structural cells, such as epithelial cells and Mast cells are major sources of TGF-β [137]. Monocytes constitutively express TGF-β1 mRNA but release the protein only when activated [138]. Pulmonary macrophages may store large amounts of TGF-β during pulmonary inflammation [139]. Role in asthma. Expression of TGF-β1 is reported to be similar in lungs from normal and asthmatic subjects. However, greater expression of TGF-β mRNA and protein by eosinophils from asthmatic subjects has been reported, with their expression correlating with the severity of asthma and the degree of subepithelial fibrosis [140]. In another study, TGF-β1 immunoreactivity was observed in the epithelium and submucosal cells, such as eosinophils and fibroblasts, but expression was greater in biopsies from patients with chronic bronchitis than in those from patients with asthma [141]. Release of TGF-β1 into bronchoalveolar lavage fluid has been observed after segmental allergen challenge [142].The possibility remains that TGF-β (together with PDGF) may be involved in the remodeling process of asthma, although it may also participate in modulating the T cell response. Nerve Growth Factor (NGF)Synthesis and release. NGF belongs to the family of neurotrophins, which control the survival, differentiation and maintenance of neurons in the peripheral and central nervous systems NGF is a homodimeric molecule. Two molecular forms of NGF exist: 7S NGF and 2.5S NGF molecules [143, 144].Many inflammatory cells express a high affinity NGF receptor: monocytes, mast cells, basophils, macrophages, T-cells and B-cells. NGF shows various effects in inflammatory models [145]. This could be relevant in relation to allergic asthma. Indeed, NGF promotes inflammatory mediator release from basophils, mast cells, T- and B- cells, eosinophils, and macrophages [146]. Role in asthma.Solar et al [147] documented a role of NGF in the induction of allergic asthma, airway hyperresponsiveness and inflammation. They also hypothesize that NGF can affect airway function by changing the properties of the sensory nerves in the airways. In order to reveal the role of NGF in asthma, they first studied the effect of NGF on airway function. They then performed a more mechanistic approach to analyze whether sensory nerve endings are involved in effects of NGF on airway function. Furthermore, they also studied the role of NGF in an allergic model of asthma. Results revealed the influence of NGF on acute bronchoconstriction induced by allergen challenge. Fibroblast growth factor (FGF)Synthesis and release. FGF represents a family of heparin-binding growth factors consisting of seven polypeptides, including acidic FGF and bFGF. Acidic FGF and bFGF are potent modulators of cell proliferation, motility, and differentiation. They are found to be associated with the extracellular matrix [148]. Role in asthma.A major role for FGF in the induction of angiogenesis has been proposed. In human adult lung, bFGF has been localized to vascular smooth muscle and endothelial cells of blood vessels of the lungs bFGF has also been detected at high levels in epithelial cells of the trachea and bronchi. bFGF increases expression of the PDGF receptor a-subunit in human airway smooth muscle and therefore indirectly stimulates proliferation [149]. Epidermal growth factor (EGF)EGF and TGF-a, which do not bind heparin, also stimulate angiogenesis. EGF expression is increased in the epithelium of patients with bronchitis and in the submucosa of patients with asthma [150]. EGF increases airway smooth muscle proliferation and ET-1 potentiates EGF-induced airway smooth muscle proliferation [151]. Increases in the number of blood vessels in asthmatic airways have been described and these growth factors may be implicated [152]. Insulin-like growth factor (IGF)IGF is produced by airway epithelial cells and is a potent mitogen for airway smooth muscle proliferation [153]. IGF activates MAP kinases in airway smooth muscle [154] and appears to mediate the proliferative effect of LTD4, at least in rabbits [155]. E. ChemokinesChemokines are small-secreted proteins and chemotactic cytokines (8 to 10 kD). Chemokines not only attract leukocytes into tissues but also regulate cell trafficking. They are classified into four subclasses: C, CC (b-chemokines), CXC (a-chemokines, C and CX3C chemokines, based on the location of the first two-cysteine residues in their sequence. To date, more than 40 chemokines are recognized [156] outof which 23 human CC chemokines, 14 human CXC, and one each of the C and CX3C chemokine subclasses have been described [152]. Chung and Barnes [8] also reported that some regulatory factors like RANTES, MCP-1, MCP-2, MCP-3, MCP-4, MCP-5 MIP-1α, eotaxin and IL-8 come under the category of Chemokines. Synthesis and release.The global biological activity of the various cytokines within each structurally related group is largely similar. Recruitment of inflammatory cells into the airway mucosa requires Th2, expression of adhesion molecules on vascular endothelium and chemokine activity. On this basis, an abundance of chemokines and chemokine receptors has been identified. The biological effects of chemokines are mediated by cell surface receptors, which belong to the superfamily of seven transmembrane G-protein coupled receptors. The CC chemokines are involved in chemoattraction of eosinophils, monocytes, and T lymphocytes and are therefore of greatest relevance in asthma [157]. The CXC or a-chemokines principally attract neutrophils and have, therefore mainly been related to acute inflammatory processes. To date, models of allergic inflammation have predominantly focused on the CC or b-chemokine family, as these express chemotactic activities towards eosinophils as well as dendritic cells, T-lymphocytes, basophils and monocytes [158]. The precise functional role of each chemokine within this group remains to fully explore. Role in asthma.There are several CXC chemokines, all of which selectively attract neutrophils. The identification of eotaxin as the first chemokine with preferential ability to recruit eosinophils has drawn much attention to this molecule [159]. Eotaxin was first discovered in the BAL fluid of guinea pigs after allergen challenge [160]. In human asthma, eotaxin is produced at high concentrations and localizes in the airway epithelium. Several other chemokines including regulated on activation, normal T-cell expressed, and secreted protein (RANTES), Monocyte chemoattractant protein (MCP)-3, and MCP-4 can also recruit eosinophils. However, it is still not clear whether all of these chemokines contribute equally to in vivo eosinophil chemotaxis, or whether a specific chemokine mediates the bulk of the chemotactic activity and can be targeted for treatment [161]. Because of its central role in eosinophil recruitment, and because of its expression on Th2cells, the CCR3 chemokine receptor might be a good target to interfere with the development of asthma. The potential role of chemokines in asthma is supported by observations that many cell types present in asthmatic airways (in particular, monocytes/ macrophages, T cells, airway smooth muscle cells, and airway epithelial cells) have the potential to generate chemokines. CC chemokines can be detected in bronchoalveolar lavage fluid, although only at low levels, even after the fluid has been concentrated. Increased levels of MCP-1, RANTES, and Macrophage inhibitory protein (MIP)-1a in asthmatic patients have been reported, and the eosinophil chemoattractant activity of bronchoalveolar lavage fluid from asthmatics were blocked by antibodies to RANTES and MCP-3 [162]. Increased expression of RANTES and MCP-3 mRNA has been reported in the airway submucosa of patients with allergic and nonallergic asthma [163]. An early report showed enhanced co-expression of IL-8 and GM-CSF in bronchial epithelial cells from patients with asthma [164]. Free IL-8 has been detected in the sera and bronchial tissue of subjects with severe atopic asthma but not in samples from normal subjects or patients with mild atopic asthma, suggesting that IL-8 may be a marker of severe asthma. IL-8 was also found to be complexedwith IgE, levels of which were increased in bronchial tissue in asthma [165]. However, in segmental local challenge studies of patients with allergic asthma, increased IL-8 levels correlated with neutrophil influx indicating that IL-8 may be mostly from patients with allergic rhinitis [166]. ConclusionThe epidemiological findings and experimental studies strongly suggest that the impact of allergens on asthma is primarily mediated by stimulation and activation of different inflammatory cells. Recently, Type-2 T-helper lymphocytes, eosinophils and mast cells are considered as the main orchestrate of allergic airway inflammation underlying asthma. The important inflammatory mediators behind asthma pathophysiology are cytokines, chemokines and leukotrienes. These mediators have the ability to initiate a cycle of events that could establish ongoing inflammation. On this basis, an understanding of cytokine biology has led to the identification of several targets that could be developed as potential treatment for asthma. Both the experimental animal models and the human genetic studies point to a limited set of cytokines and cytokine receptors as crucial targets for the treatment of asthma, particularly IL-2, IL-4, IL-5, IL-9, IL-13, chemokines and leukotrienes and their respective receptors. Which cytokine is the best target remains to be determined, but it is tempting to speculate that the heterogeneity of asthma might reflect heterogeneity of mechanisms and cytokines involved. Indeed, there is currently a large drive to test several inhibitors of cytokine effects in asthma, such as inhibitors of IL-5 and IL-9, which is a cytokine central to the development of tissue eosinophilic inflammation. Clinical studies of an antibody to IL-5 and IL-9 are currently underway. However, it is unclear to what extent blocking the effects of one single cytokine out of all the cytokines that are potentially involved will lead to important therapeutic effects. A study of the actions of these cytokines may provide other ways of blocking the effects of groups of cytokines. Corticosteroids may have beneficial effects in asthma by their ability to block the generation of several pro-inflammatory cytokines. Therefore, not only will greater understanding of asthma pathogenesis be achieved by studying cytokine biology, but also the potential for newer more effective treatments, perhaps even a cure. Currently, in vivo animal models have confirmed the role of T-helper type-2 lymphocytes in airway inflammation. Th2 cells seem to be the main factor in determining the degree of airway inflammation and hyperresponsiveness. It would appear that cytokine production, rather than the influx of eosinophils or production of immunoglobulin-E, is the cause of bronchial hyperresponsiveness. Importantly, biopsy studies clearly illustrate the presence of a complex cytokine network in asthmatic airways. The redundancy of and mutual interaction between the cytokines within this network needs to be borne in mind when trying to asses the functional role of an individual cytokine in the pathophysiology of asthma. Neutralizing antibodies as well as soluble receptors may also offer an opportunity to inhibit selectively asthmatic inflammation without major side effects. However by inhibiting these cytokines not only asthmatic inflammation is decreased but other important regulatory functions of cytokines are also abolished. Studies of airways in chronic asthmatics by bronchoscopic methods and induced sputum have provided much helpful and insightfuldata. Experiments such as the bronchial challenge with allergenprovide valuable insights into the allergic inflammatory responsebut no scientific reports shows the actual remodeling process. Thus, the process of airway wall remodelingis still not understood and requires investigation into its mechanismsand the role of drugs in its reversal and prevention. The genetic approaches described above have provided wealth of valuable information but also have important limitations. In addition, simply knowing genes are associated with asthma will not reveal everything about the development of the disease or the value of the gene/gene product as therapeutic target. Pharmacogenetic studies will offer a better understanding of full or partial resistance to treatment and progression of more or less severe asthma phenotype. Such molecular genetic studies may open a way for better patient management. Lastly, we would like to convey our message for younger research scientist, the evaluation of new therapeutic agents is the today’s need for managing asthma and related respiratory disorders. It should include investigation of the effects of drugs especially on airway inflammation, remodeling, hyperresponsiveness and also infiltration and activation of eosinophils. References
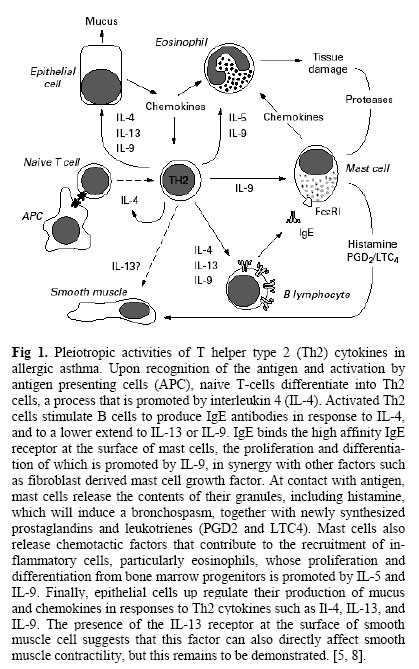
Copyright © 2005 by Razi Institute for Drug Research (RIDR) The following images related to this document are available:Photo images[pt06001f1.jpg] | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||
{kind=link}